Pratiquement tous les transferts d’électrons hors équilibre sur Terre sont pilotés par un ensemble de machines nanobiologiques composées en grande partie de complexes protéiques multimériques associés à un petit nombre de groupes prothétiques. Ces machines ont évolué exclusivement chez les microbes au début de l’histoire de notre planète mais, malgré leur ancienneté, elles sont hautement conservées. Par conséquent, bien qu’il existe une énorme diversité génétique dans la nature, il reste un ensemble relativement stable de gènes fondamentaux codant pour les principales réactions redox essentielles à la vie et aux cycles biogéochimiques. Ces gènes ont été créés et ont coévolué avec des cycles biogéochimiques et ont été transmis de microbe à microbe principalement par transfert horizontal de gènes. Un défi majeur dans les décennies à venir est de comprendre comment ces machines ont évolué, comment elles fonctionnent, et les processus qui contrôlent leur activité à l’échelle moléculaire et planétaire. https://www.science.org/doi/10.1126/science.1153213
Nous avons constaté qu’une diminution de la diversité microbienne (i) affectait la décomposition des sources de carbone autochtones et allochtones, réduisant ainsi les émissions mondiales de CO2
jusqu’à 40 %, et (ii) façonnait la source de CO2 émission vers une décomposition préférentielle de la plupart des sources de C dégradables. Nos résultats ont également révélé que l’importance de l’effet de diversité augmente avec la disponibilité des nutriments. Dans l’ensemble, ces résultats montrent que le cycle du carbone dans le sol pourrait être plus vulnérable aux changements de diversité microbienne que prévu par les études précédentes, en particulier dans les écosystèmes exposés aux apports de nutriments. Ainsi, les préoccupations concernant la préservation de la diversité microbienne peuvent être très pertinentes dans le contexte actuel de changement global supposé avoir un impact sur la biodiversité des sols et sur les apports pulsés de résidus végétaux et de rhizodépôts dans le sol.
IMPORTANCE Avec des centaines de milliers de taxons par gramme de sol, la diversité microbienne domine la biodiversité des sols. Bien que de nombreuses études aient établi que les communautés microbiennes réagissent rapidement aux changements environnementaux, la relation entre la diversité microbienne et le fonctionnement du sol reste controversée. https://pubmed.ncbi.nlm.nih.gov/29453268/
Qu’est ce que l’eau ?
L‘eau structurée a des propriétés différentes de l’eau classique. L’eau est le support des molécules les plus importantes de la vie, comme les protéines et l’ADN. Dans les cellules, les gels et le moteur de la vie, il est prouvé que l’eau est absolument nécessaire à toutes les activités des cellules. L’eau de nos cellules n’est pas comme l’eau d’un verre. Elle est en fait presque aussi ordonnée qu’un cristal. Comme la glace, elle piège les particules et les solutés lorsqu’elle se forme. L’espace qui en résulte est appelé la zone d’exclusion (ZE). Le Dr Pollack a découvert une nouvelle phase de l’eau. L’eau primaire est H2O, mais cette nouvelle phase aqueuse, l’eau structurée dans la zone d’exclusion, est H3O2. C’est la phase aqueuse nouvellement découverte. Si vous comptez la quantité d’hydrogène et d’oxygène, vous constaterez que ce n’est pas H2O. Si le quatrième état d’eau est appelée zone d’exclusion ou EZ, c’est parce que l’équipe du Dr Pollack a d’abord constaté qu’elle était tout à fait unique. Même les petites molécules sont exclues de l’eau structurée. Étonnamment, l’eau structurée est très abondante, y compris dans la plupart de vos cellules. Même vos tissus extracellulaires sont remplis de cette eau. Eau de source – Formée sous pression (en profondeur). Fonte des glaciers – Lorsque la glace fond, elle devient de l’eau structurée (eau EZ). La phase entre le liquide et le solide est de l’eau structurée.
- Formation de courants de Foucault – Les courants de Foucault se produisent naturellement dans la nature comme les ruisseaux, les rivières, les chutes d’eau, etc. Les courants de Foucault sont des perturbations ou des mouvements mécaniques.
Les tourbillons ou vortex sont un moyen très efficace d’ajouter de la texture. Il existe des dispositifs de tourbillonnement de l’eau sur le marché. L’un de ces appareils est une unité d’eau structurée d’apparence naturelle. - Le jus – est l’écoulement de l’eau des cellules végétales. De l’eau de jus structurée !
- Antioxydants – La plupart des tissus de notre corps sont négatifs. Nos cellules sont chargées négativement ; les oxydants sont chargés positivement. Les antioxydants maintiennent la charge négative de notre corps.
- La lumière du soleil – est vitale pour notre santé. La lumière crée de l’eau structurée (eau EZ).
- Circulation – Les globules rouges voyagent dans les capillaires ; la lumière est ce qui alimente la rivière. Ajoutez de la lumière et le flux augmente. Un élément autre que le cœur (la pression) fait circuler le sang.
- Infrarouge – l’énergie est partout. Elle alimente les processus de votre corps.
La quatrième phase d’eau : à partir de ce que nous savons de l’eau. . . par une expérience simple, nous avons découvert la quatrième phase de l’eau. Quelle est la nature de cette quatrième phase de l’eau ? Pourquoi c’est intéressant Cela fonctionne pour tout ce qui entre en contact avec l’eau. On la trouve dans le ciel et les nuages. On la trouve dans les océans, les lacs et les rivières et elle remplit l’intérieur de notre corps.

Effet du tensioactif soluble sur la persistance des bulles et des particules d’aérosol produites par les bulles
https://agupubs.onlinelibrary.wiley.com/doi/full/10.1002/jgrd.50186 [9]
L’effet du tensioactif soluble sur la persistance des bulles d’eau salée et leur capacité à produire des particules d’aérosol lors de leur éclatement a été étudié. Des ensembles de bulles individuelles millimétriques ont été produits dans des solutions de NaCl de différentes concentrations de tensioactif. L’efficacité de la production d’aérosols – une propriété fondamentale des bulles simples définie comme le nombre de particules produites par surface de film à bulles a diminué de 79 % à 98 % suite à l’ajout d’un tensioactif et à l’augmentation de la pression du film de solution de 1 à 2 à 7 à 27 mN. m −1 . La persistance des bulles à la surface de l’eau et l’épaisseur de leur pellicule ont été étudiées par vidéographie à haute vitesse. L’ajout d’un tensioactif soluble a augmenté la persistance moyenne des bulles, ce qui a donné plus de temps aux bulles pour s’écouler et s’éclaircir à l’aide de flux de régénération marginaux. Les épaisseurs du capuchon du film à bulles variaient d’environ 1 µm pour les bulles relativement propres et de courte durée à moins de 0,1 µm pour les bulles persistantes stabilisées par un tensioactif. La suppression de la production d’aérosols à partir des bulles stabilisées par un tensioactif peut résulter de l’amincissement spectaculaire de leurs coiffes ou de la réduction des forces de surface à une pression de film élevée. Les fonctions de source d’aérosols Sea Spray précédemment rapportées ont été comparées aux efficacités de production d’aérosols mesurées et se sont révélées être d’une ampleur nettement supérieure, ce qui suggère qu’il existe une source de particules provenant des whitecaps qui n’a pas été capturée dans ces expériences à bulle unique.
- L’efficacité de la production d’aérosols a fortement diminué avec l’augmentation de la pression du film
- Les bulles stabilisées par un tensioactif persistaient plus longtemps et avaient des capsules de film beaucoup plus fines
Microcouches de surface de la mer : une perspective physico-chimique et biologique unifiée de l’interface air-océan
https://www.sciencedirect.com/science/article/abs/pii/S0079661112000924 [8]
La microcouche de surface de la mer (SML) couvre plus de 70 % de la surface terrestre et constitue l’interface de couche limite entre l’océan et l’atmosphère. Cet important système biogéochimique et écologique est essentiel à un large éventail de processus du système terrestre, notamment la synthèse, la transformation et le cycle des matières organiques, ainsi que l’échange air-mer de gaz, de particules et d’aérosols. Dans cette revue, nous discutons du paradigme SML, en tenant compte des caractéristiques physico-chimiques et biologiques qui définissent la structure et la fonction du SML. Ceux-ci incluent des enrichissements en molécules biogéniques telles que les glucides, les lipides et les matières protéiques qui contribuent au cycle du carbone organique, des assemblages microbiens distincts qui participent aux échanges gazeux air-mer, la génération d’aérosols actifs sur le climat et l’accumulation de polluants anthropiques avec des implications potentiellement graves. pour la santé de l’océan. Des gradients physiques, chimiques et biologiques typiquement importants séparent ainsi le SML de l’eau sous-jacente et les preuves disponibles impliquent que le SML conserve son intégrité dans des conditions environnementales très variées. Une gamme de polluants chimiques se trouvent dans la SML, notamment des hydrocarbures, des composés organochlorés et des métaux traces, qui peuvent généralement être dix à cent fois plus concentrés que dans les eaux sous-jacentes. Les polluants SML présentent donc toute une série de préoccupations environnementales et écotoxicologiques potentielles pour le fonctionnement de l’écosystème SML. Notamment, des accumulations plus élevées se trouvent dans le SML des eaux côtières proches des sources anthropiques de pollution.
Effets Marangoni induits par les tensioactifs générés par le plancton
https://www.sciencedirect.com/science/article/abs/pii/S0924796305000631 [7]
L’intérêt de créer une capacité de surveillance des surfactants océaniques est dû à l’hypothèse selon laquelle la prévalence des films océaniques suit de près la productivité biologique. La répartition mondiale actuelle des tensioactifs capables d’affecter les processus d’échange est largement inconnue, tout comme les facteurs induisant les répartitions futures. On pense que la plus grande source de tensioactifs à la surface de la mer est la production d’organismes marins, principalement le phytoplancton, qui exsudent des tensioactifs naturels sous forme de sous-produits métaboliques. Les tensioactifs relativement solubles produits par le phytoplancton peuvent fortement affecter la dissipation des turbulences proches de la surface et conduire à des modifications des processus d’échange gazeux air-mer. En plus de ces effets, la présence de tensioactifs à la surface de la mer pourrait également induire des mouvements dus aux changements de tension superficielle. Il est en effet bien connu que la présence de tensioactifs réduit la tension superficielle à l’interface air/eau. Le gradient de tension superficielle qui en résulte conduit à une contrainte de cisaillement superficielle, communément appelée contrainte de Marangoni, qui, en raison de la viscosité, influence l’écoulement global qui est accéléré ou retardé, en fonction de l’orientation d’un tel gradient. Il est donc raisonnable de s’attendre à ce que les tensioactifs générés par le phytoplancton puissent provoquer ce type d’effets thermocapillaires sur la circulation de l’eau.
Composition en matière organique et enrichissement en polluants de la microcouche de surface de la mer à l’intérieur et à l’extérieur des nappes
https://www.sciencedirect.com/science/article/abs/pii/014111369390100E [6]
Des échantillons de microcouches de surface ont été collectés à l’aide d’un dispositif rotatif dans huit stations situées dans une zone côtière et examinés, avec leurs eaux en vrac associées, pour déterminer leur composition en matière organique. À l’exception des pigments, tous les groupes de composés examinés, en particulier les hydrocarbures et les détergents anioniques, présentent une accumulation dans la microcouche superficielle ; l’enrichissement est plus élevé dans les échantillons collectés à l’intérieur des nappes qu’à l’extérieur. Une diminution des mesures de tension superficielle est liée à l’accumulation de matière organique dans les nappes. Dans les microcouches superficielles, la majeure partie de la fraction organique identifiée est constituée de glucides et de composés protéiques plutôt que de lipides. D’après ce que l’on peut déduire d’une estimation des lipides, des sucres et des protéines-carbone, la moitié du poids de carbone organique particulaire enregistré est d’une forme complexe non identifiée. L’étude de la cinétique d’hydrolyse de la fraction particulaire glucidique montre que la matière organique se présente sous une forme moins condensée dans les microcouches superficielles que dans les échantillons d’eau massifs. Une relation étroite apparaît entre l’accumulation de polluants et la baisse de la tension superficielle des nappes. Cela a une conséquence directe : l’effet amortisseur des vagues qui lisse la surface de la mer, rendant les nappes observables, caractéristiques de l’accumulation de polluants.
Répartition des organochlorés dans la phase dissoute et en suspension de la microcouche de surface de la mer et de l’eau de mer à Hong Kong, Chine
https://www.sciencedirect.com/science/article/abs/pii/S0025326X05005564 [2]
La microcouche de surface (SML) de l’océan représente l’interface entre l’atmosphère et le corps de la surface de l’océan et a une épaisseur typique de 40 à 100 μm. Le SML est enrichi de composés organiques naturels, notamment de protéines, de lipides et de tensioactifs organiques, ce qui lui confère une composition chimique distincte. On sait que les contaminants anthropiques, y compris les polluants organiques persistants (POP), s’hyper-accumulent dans le SML, entraînant un enrichissement relatif par rapport à la colonne d’eau sous-jacente. Il existe également des preuves selon lesquelles les particules floculées, flottant à la surface de la mer, peuvent agir comme piégeurs de POP dans la colonne d’eau des zones côtières. Les niveaux de concentration et les facteurs d’enrichissement des POP dans les SML échantillonnés dans différentes parties du monde. Le SML est considéré comme un écosystème unique avec une grande biodiversité d’organismes et constitue un habitat essentiel pour les premiers stades de vie de nombreuses espèces de poissons et d’invertébrés. Il a été démontré que les POP présents dans les SML des eaux côtières polluées induisent une réaction toxique chez les larves de poissons. En tant que tel, l’enrichissement en POP dans le SML représente une menace potentielle à la fois pour la biodiversité marine naturelle et pour la pêche commerciale.
Caractériser la contamination de la surface des océans : composition, épaisseur du film et rhéologie
https://www.sciencedirect.com/science/article/abs/pii/S0025326X22009699 [1]
La pollution marine est une source d’inquiétude pour l’humanité depuis plusieurs décennies en raison des graves menaces qu’elle fait peser sur l’écosystème. En plus de nuire à la vie aquatique, les polluants pénètrent également dans la chaîne alimentaire et finissent par atteindre le corps humain. Les déchets marins comprennent les substances qui flottent à la surface de l’eau formant une couverture, ou celles qui coulent dans l’océan. Un exemple classique de contamination de surface est la zone de déchets du Grand Pacifique, qui est une étendue isolée de débris comprenant des plastiques, couvrant une superficie d’environ 1,6 million de kilomètres carrés de surface de l’eau. Bien qu’il s’agisse du plus grand cas connu de contamination de surface, il existe d’autres cas importants, comme celui du dépotoir de l’océan Indien. Bien que le plastique constitue une grande partie des déchets marins formant ces dépôts de déchets, il existe d’autres substances qui contribuent à la contamination. La pollution des océans est due à de nombreuses sources, notamment les déversements/accidents de pétrole, les matières mortes en décomposition, les biofilms, les effluents industriels, les eaux usées provenant du ruissellement terrestre, le transport atmosphérique, l’acidification des océans, etc.
En plus de provoquer des changements dans l’écosystème marin, la contamination des surfaces est également connue pour affecter des phénomènes naturels tels que les vagues, qui influencent l’échange de masse, de quantité de mouvement et d’énergie à la frontière commune partagée par l’océan et l’atmosphère. L’une des conséquences majeures de la contamination des surfaces océaniques est l’amélioration de l’amortissement des vagues. L’amortissement des vagues dû à la contamination de la surface a été observé depuis l’époque de Pline l’Ancien, lorsqu’il était courant pour les voyageurs de déverser intentionnellement du pétrole à la surface de la mer pour « calmer » les vagues. Depuis le début du XXe siècle, il y a eu un intérêt croissant pour l’étude de l’amortissement des vagues dû aux tensioactifs. La plupart des résultats concernent des ondes capillaires-gravitaires à l’interface contaminée de deux fluides visqueux. Par exemple, les résultats sont nombreux sur la dissipation des vagues de houle, connues pour affecter le climat vent-vagues. La présence de tensioactifs à la surface de l’eau facilite également les ondes de dilatation, qui sont créées lorsque les forces de Marangoni sont produites en raison de la non-uniformité spatiale de la tension interfaciale résultant de la contrainte de l’interface due à l’action des ondes transversales.
Influence du non ionique Tensioactif sur la formation de biofilm attaché et la biodisponibilité du phénanthrène lors de simulations Tensioactif Bioremédiation améliorée
https://pubs.acs.org/doi/10.1021/es0701154 [14]
Tensioactif : les effets sur la formation de biofilms préformés attachés et la biodégradation du phénanthrène sorbé dans le sol ont été surveillés à l’aide d’un système de cellules à circulation à écoulement continu. Les résultats obtenus dans cette étude révèlent que la formation de biofilm bactérien avec des substances polymères extracellulaires était la stratégie des bactéries pour utiliser des HAP partiellement solubles. Cependant, en présence de tensioactif, cette stratégie a été modifiée. La présence de tensioactif à toutes les concentrations, les aspects physiologiques du biofilm attaché ont changé et la biodisponibilité du phénanthrène a augmenté avec l’ajout de tensioactif au dessus du cmc.
Réactivité et devenir des tensioactifs synthétiques en milieux aquatiques
https://www.sciencedirect.com/science/article/abs/pii/S016599360800112X [15]
Depuis la seconde moitié du XXe siècle, la production et la consommation de composés organiques ont connu une croissance exponentielle dans le monde. Environ 1 500 nouveaux composés sont introduits sur le marché chaque année, selon l’Agence européenne pour l’environnement (AEE). Pour la plupart de ces substances, la plupart des aspects de leur comportement environnemental sont totalement inconnus, tout comme leur impact possible sur les écosystèmes une fois rejetées après utilisation.
Parmi ces composés, les tensioactifs synthétiques sont parmi les plus importants. La demande de ces produits chimiques en 2003 était estimée à plus de 9 millions de tonnes, ils présentent donc un intérêt particulier. Des efforts considérables ont été consacrés à leur détermination et à leur surveillance, tant par des tests en laboratoire que dans l’environnement, notamment dans les systèmes aquatiques, où ils sont introduits après leur rejet des stations d’épuration des eaux usées (STEP) et la valorisation agricole des boues provenant des STEP.
Les tensioactifs sont utilisés dans une grande variété d’applications, principalement dans la formulation de détergents, mais également comme ingrédients dans des produits de soins personnels, des peintures, des pesticides et de nombreux autres produits. En conséquence, de nombreux types différents de tensioactifs ont été synthétisés, bien qu’ils puissent être classés en quatre groupes différents en fonction de leur charge. La plupart d’entre eux appartiennent aux deux principaux groupes, les anioniques et les non ioniques, qui représentent plus de 90 % de la production européenne. Concernant les anioniques, les alkylbenzènesulfonates linéaires (LAS) sont les tensioactifs les plus utilisés, avec une production annuelle totale de 434 000 tonnes rien qu’en Europe, selon les données du CESIO (Comité Européen des Agents de Surface et de leurs Intermédiaires Organiques). Ils sont couramment utilisés dans les détergents ménagers et les nettoyants tout usage.
Les éthoxysulfates d’alkyle (AES) et les sulfates d’alkyle (AS), qui sont principalement utilisés comme ingrédients dans les shampooings et autres produits de soins personnels en raison de leurs excellentes propriétés moussantes, arrivent en deuxième position en termes de production, avec une production combinée de 404 000 tonnes.
Parmi les tensioactifs non ioniques, les polyéthoxylates d’alcool (OEA) sont actuellement produits en plus grand volume, 747 000 tonnes en Europe en 2000, et sont utilisés dans les nettoyants domestiques et industriels. En deuxième position en volume se trouvent les éthoxylates d’alkylphénol (APEO), bien que ces composés aient connu une diminution significative de leur volume de production ces dernières années en raison de politiques environnementales restrictives, visant à prévenir les effets négatifs, tels que l’œstrogénicité, manifestés par leurs intermédiaires de dégradation. .
Pour plusieurs raisons, il est compliqué et fastidieux d’identifier et de surveiller les niveaux de tensioactifs une fois qu’ils atteignent les milieux aquatiques, ainsi que d’étudier leur comportement et leur devenir dans ces systèmes. Premièrement, les tensioactifs sont souvent vendus sous forme de mélanges commerciaux, qui peuvent comprendre des centaines d’homologues, d’isomères et/ou d’éthoxymères différents, chacun présentant des propriétés physico-chimiques qui peuvent différer considérablement les unes des autres (par exemple, le log K oe calculé pour les AEO est de 2,67). –6,69, selon l’homologue ou l’éthoxymère). La séparation et la quantification des composants de ces mélanges sont donc nécessaires pour une meilleure compréhension du comportement environnemental des tensioactifs. Cela nécessite le développement de méthodologies analytiques puissantes, la plupart basées sur l’utilisation de la chromatographie en phase gazeuse (GC) ou de la chromatographie liquide (LC) et de la spectrométrie de masse (MS), comme nous l’expliquons ci-dessous. De plus, l’analyse des tensioactifs dans les matrices environnementales présente un défi supplémentaire, en particulier lorsqu’il s’agit d’échantillons solides (par exemple, sédiments et matières en suspension), mais également d’échantillons aqueux, car les composés cibles ont tendance à être présents en dessous du niveau du ppb. Ainsi, des protocoles fiables d’extraction, de purification et de préconcentration doivent être développés pour les échantillons solides et aqueux afin d’éliminer autant d’interférences que possible sans sacrifier les valeurs de récupération élevées des tensioactifs et de leurs métabolites.
La connaissance de la réactivité de ces composés dans les systèmes aquatiques peut être partiellement déduite des données d’échantillonnage sur le terrain disponibles. La dégradation et la sorption ont ainsi été identifiées comme les deux principaux processus responsables de l’élimination des tensioactifs non seulement après, mais aussi avant, qu’ils atteignent les milieux aquatiques. Une fois utilisés, ces produits chimiques sont éliminés dans les égouts, où on estime que 50 % en volume sont dégradés, 25 % étant adsorbés dans les matières en suspension et 25 % dissous. Plus tard, les tensioactifs atteignent les STEP, où 95 à 99 % sont généralement éliminés lors du traitement.
La plupart des tensioactifs utilisés sont biodégradés en aérobie lors du traitement secondaire, mais une fraction considérable est également éliminée sous forme de boues (15 à 37 % dans le cas des LAS à plus de 90 % pour les nonylphénols (NP) les plus hydrophobes). Ces boues, souvent utilisées en agriculture après une digestion anaérobie préalable, sont également une source potentielle de contamination des sols, des eaux souterraines et des rivières adjacentes, car elles ont tendance à contenir des concentrations relativement importantes de tensioactifs (plusieurs g/kg), entre autres contaminants.
Enfin, une fois que les tensioactifs atteignent la colonne d’eau, ils se dégradent relativement rapidement ; les demi-vies estimées sont de plusieurs heures à quelques jours, selon le surfactant et les facteurs environnementaux (par exemple, température et salinité). Cependant, des proportions importantes (de l’ordre de 10 % pour les LAS et AES à plus de 50 % pour les APEO et AEO) sont fixées sur les matières en suspension et finissent dans les sédiments, où, en dessous de quelques cm de profondeur, règnent des conditions anoxiques. , les tensioactifs ne peuvent donc être dégradés que par des voies anaérobies lentes et ont tendance à être préservés le long de la colonne sédimentaire.
Compte tenu de tout cela, le comportement des tensioactifs ne peut être pleinement compris qu’après une caractérisation appropriée des processus de sorption et de dégradation en laboratoire dans des conditions contrôlées. De cette manière, l’influence de ces processus peut être examinée de manière adéquate et plusieurs paramètres (par exemple, les coefficients de sorption, les demi-vies ou les facteurs de bioconcentration) peuvent être calculés pour améliorer l’évaluation des risques liés à ces composés.
Deux régimes de dégradation de la mousse blanche en laboratoire : contrôlé par un panache de bulles et stabilisé par un tensioactif
https://journals.ametsoc.org/view/journals/phoc/43/6/jpo-d-12-0148.1.xml?alreadyAuthRedirecting [3]
Cela est vrai, que des tensioactifs soient présents ou non. Cependant, en présence de tensioactifs, la mousse blanche est stabilisée et persiste environ 3 fois sa valeur dans l’eau de mer propre. Nous explorons les rôles que jouent la flottabilité des bulles et le tensioactif soluble dans la détermination du temps de décomposition de la mousse à tête blanche générée à partir de paquets d’ondes focalisés dans le canal à vagues du canal de verre de la Scripps Institution of Oceanography. La tension superficielle a été modifiée par l’ajout du tensioactif soluble Triton X-100 (204 µg L -1 ). Cela a abouti à une tension superficielle de 70 mN m −1 (±0,5 mN m −1 ), ce qui a donné une pression de film d’environ 4 mN m −1 . La pression du film peut être définie comme la différence de tension superficielle entre l’eau de mer non contaminée (nominalement 74 mN·m −1 ) et l’eau de mer contaminée. Nous notons que, malgré l’implication d’une eau exempte de tout matériau tensioactif, l’eau de mer filtrée contenait probablement de faibles niveaux de tensioactif. La pression du film mesurée pour le régime d’eau propre était de 1 mN m −1 (±0,5 mN m −1 ), indiquant des conditions d’eau propre acceptables pour nos besoins. En effet, notent qu’il est « extrêmement difficile » d’éliminer les tensioactifs même lors de la production d’eau propre pour des expériences en laboratoire. Les deux groupes de données montrent une corrélation entre le temps de décroissance de la mousse et du panache, qui augmente avec l’augmentation de la pente des vagues. Les figures illustrent respectivement la relation entre la pente des vagues et l’échelle de temps de désintégration de la mousse, et la pente des vagues et l’échelle de temps de désintégration du panache.
Microcouches de surface de la mer : une perspective physico-chimique et biologique unifiée de l’interface air-océan
https://www.sciencedirect.com/science/article/abs/pii/S0079661112000924 [4]
La microcouche de surface de la mer (SML) couvre plus de 70 % de la surface terrestre et constitue l’interface de couche limite entre l’océan et l’atmosphère. Cet important système biogéochimique et écologique est essentiel à un large éventail de processus du système terrestre, notamment la synthèse, la transformation et le cycle des matières organiques, ainsi que l’échange air-mer de gaz, de particules et d’aérosols. Dans cette revue, nous discutons du paradigme SML, en tenant compte des caractéristiques physico-chimiques et biologiques qui définissent la structure et la fonction du SML. Ceux-ci incluent des enrichissements en molécules biogéniques telles que les glucides, les lipides et les matières protéiques qui contribuent au cycle du carbone organique, des assemblages microbiens distincts qui participent aux échanges gazeux air-mer, la génération d’aérosols actifs sur le climat et l’accumulation de polluants anthropiques avec des implications potentiellement graves pour la santé de l’océan. Des gradients physiques, chimiques et biologiques typiquement importants séparent ainsi le SML de l’eau sous-jacente et les preuves disponibles impliquent que le SML conserve son intégrité dans des conditions environnementales très variées. À l’appui de cela, nous présentons des données chronologiques inédites sur la composition du bactérioneuston et l’activité du tensioactif SML immédiatement après la perturbation physique du SML ; ceux-ci impliquent des délais de l’ordre de quelques minutes pour le rétablissement du SML suite à une perturbation. Une approche progressive de la compréhension du SML et donc de son rôle dans la biogéochimie mondiale ne peut être réalisée qu’en considérant comme un tout intégré tous les composants clés de cet environnement complexe.
La microcouche de surface de la mer (SML) constitue les dizaines à centaines de μm supérieurs de la surface de l’océan qui est en contact direct avec l’atmosphère et possède des propriétés physicochimiques et biologiques mesurables distinctes de celles des eaux sous-jacentes . Les preuves de l’existence du SML ont une longue histoire. Les nappes naturelles à la surface de la mer dans les zones de forte activité biologique, comme dans les eaux côtières, sont bien documentées. Les travaux menés dans la mer des Sargasses ont établi un lien précoce entre les nappes visibles et les proliférations de phytoplancton, et ont montré que les nappes peuvent avoir des effets fondamentaux sur les propriétés physiques de la surface de la mer. Des preuves d’une origine et d’une activité biologiques ont été révélées dans les enrichissements en glucides des nappes par rapport aux eaux sous-jacentes.
Nous discutons du rôle central des micro-organismes dans le maintien des propriétés physicochimiques du SML et dans le maintien des processus biogéochimiques clés. Les dernières sections se concentrent sur trois processus spécifiques : le transfert de gaz air-mer, la production d’aérosols et la pollution. L’étude de la vie microbienne dans les films de surface aquatiques n’est pas nouvelle. Le botaniste suédois Einar Naumann a introduit en 1917 le terme neuston pour identifier les organismes associés à l’interface air-eau qui étaient écologiquement distincts du plancton dans l’eau sous-jacentes. Par la suite, des études ont montré que la diversité des micro-organismes dans le SML peut différer considérablement de celle de l’eau sous-jacente. Les échanges gazeux air-mer jouent un rôle fondamental dans la biogéochimie mondiale et sont régis par une diffusion moléculaire et turbulente. La diffusion turbulente est définie par les échelles de longueur et de vitesse des tourbillons turbulents dans l’eau en vrac et dans l’air proche de l’interface air-eau, tandis que la diffusion moléculaire opère à l’échelle du SML où ces tourbillons sont supprimés. Une gamme de polluants chimiques se trouvent dans la SML, notamment des hydrocarbures, des composés organochlorés et des métaux traces, qui peuvent généralement être dix à cent fois plus concentrés que dans les eaux sous-jacentes. Les polluants SML présentent donc toute une série de préoccupations environnementales et écotoxicologiques potentielles pour le fonctionnement de l’écosystème SML (Wurl et Obbard, 2004). Notamment, des accumulations plus élevées se trouvent dans le SML des eaux côtières proches des sources anthropiques de pollution.
Modélisation numérique des effets Marangoni induits par les tensioactifs générés par le plancton
https://www.sciencedirect.com/science/article/abs/pii/S0045653512011046 [6]
Au cours des dernières années, la microcouche de surface de la mer et son importance relative dans le contexte des processus environnementaux marins et du changement global ont suscité un intérêt croissant. Les substances tensioactives naturelles se trouvent souvent en grandes quantités à la surface de la mer, et il existe de plus en plus de preuves de l’importance des films de surface dans le transfert de masse, de chaleur et de quantité de mouvement à travers l’interface air-mer.
L’accumulation de produits chimiques organiques naturels modifie les propriétés physiques et optiques de la surface de la mer. Les tourbillons océaniques deviennent visibles sur les images optiques et SAR (Synthetic Aperture Radar) lorsque la surface de l’océan est partiellement recouverte de films de surface, c’est-à-dire lorsque le courant de surface associé à un tourbillon a entraîné des nappes naturelles de surface. L’intérêt de créer une capacité de surveillance des surfactants océaniques est dû à l’hypothèse selon laquelle la prévalence des films océaniques suit de près la productivité biologique. La répartition mondiale actuelle des tensioactifs capables d’affecter les processus d’échange est largement inconnue, tout comme les facteurs induisant les répartitions futures.
On pense que la plus grande source de tensioactifs à la surface de la mer est la production d’organismes marins, principalement le phytoplancton, qui exsudent des tensioactifs naturels sous forme de sous-produits métaboliques. Les tensioactifs relativement solubles produits par le phytoplancton peuvent fortement affecter la dissipation des turbulences proches de la surface et conduire à des modifications des processus d’échange gazeux air-mer. En plus de ces effets, la présence de tensioactifs à la surface de la mer pourrait également induire des mouvements dus aux changements de tension superficielle.
Il est raisonnable de s’attendre à des valeurs plus significatives de la contrainte de Marangoni là où se produisent de fortes variations de la concentration de matières tensioactives, comme par exemple dans les régions côtières d’upwelling, dans les estuaires et dans les zones d’écoulements convergents en surface, où de fortes des variations horizontales de la répartition du phytoplancton, et donc de la concentration en tensioactifs, apparaissent. Une circulation renversée provoque une augmentation locale de la biomasse phytoplanctonique, avec une libération de tensioactifs qui migrent vers la surface. Les variations horizontales de tension superficielle dues aux tensioactifs générés par le phytoplancton entraînent un stress de Marangoni en surface.
La microcouche de surface de la mer : Biologie, chimie et enrichissement anthropique
https://www.sciencedirect.com/science/article/abs/pii/0079661182900015 [11]
Des études récentes soulignent de plus en plus l’interface entre l’atmosphère mondiale et l’hydrosphère (la microcouche de la surface de la mer) comme un habitat biologique important et un point de collecte de matières anthropiques. Les techniques d’échantillonnage nouvellement développées collectent différentes fractions qualitatives et quantitatives de la surface supérieure de la mer à des profondeurs allant de moins d’un micron à plusieurs centimètres.
La microcouche fournit un habitat à un biote, notamment aux larves de nombreuses espèces de pêche commerciale, dont la densité est souvent très enrichie par rapport aux eaux souterraines situées à seulement quelques centimètres en dessous. Les enrichissements courants en bactérioneuston, phytoneuston et zooneuston sont respectivement 10 2 −10 4 , 1−10 2 et 1−10. Les relations trophiques ou le fonctionnement intégré de ces communautés neustoniques n’ont pas été examinés.
Les forces de tension superficielle fournissent une microcouche physiquement stable, mais soumise à des variations environnementales et climatiques plus importantes que la colonne d’eau. Un certain nombre de processus physiques mal compris contrôlent le mouvement et le flux des matériaux à l’intérieur et à travers la microcouche. La microcouche est généralement recouverte d’un film organique naturel de lipides et d’acides gras recouvrant un complexe protéique polysaccharide.
La microcouche sert à la fois de source et de puits pour les matériaux présents dans l’atmosphère et la colonne d’eau. Parmi ces matériaux se trouvent de grandes quantités de substances anthropiques qui sont fréquemment présentes à des concentrations 10 2 −10 4 supérieures à celles de la colonne d’eau. Ceux-ci comprennent les plastiques, les morceaux de goudron, les hydrocarbures polyaromatiques, les hydrocarbures chlorés et les métaux potentiellement toxiques, tels que le plomb, le cuivre, le zinc et le nickel. La manière dont les processus uniques se produisant dans la microcouche affectent le sort des substances anthropiques n’est pas encore claire.
Propriétés chimiques de surface des nappes marines en bandes
https://www.sciencedirect.com/science/article/abs/pii/0011747174900229 [10]
Les concentrations en surface et les pressions de film des constituants organiques formant la nappe ont été mesurées in situ et sur des échantillons d’eau de surface de l’océan Pacifique, à l’ouest de San Diego. Les données de tension superficielle, précises à ±0,6 dyne/cm, ont indiqué que les pressions de film variaient aussi bas que quelques dynes/cm dans les nappes rubanées associées aux ondes internes. Cette observation est cohérente avec les données précédemment rapportées qui montrent que les films de surface insolubles dans l’eau amortissent fortement les ondes capillaires à des pressions de film aussi basses que 0,5 à 1,0 dyne/cm. Une certaine matière tensioactive était présente dans tous les échantillons, qu’ils proviennent d’eau ondulée « propre » ou de nappes visibles. Les pressions du film ont été mesurées en fonction de la concentration superficielle des constituants filmogènes. Ces données indiquaient que les concentrations dans de nombreuses zones coulées étaient juste suffisantes pour amortir ou résister à la formation d’ondulations. Cette observation a également été corroborée par les données de tension superficielle in situ . Il a été estimé qu’une compression quadruple de la couche superficielle par des forces convergentes horizontales était nécessaire pour créer des nappes visibles à cet endroit.
De nombreuses informations sont désormais disponibles démontrant que certains films monomoléculaires peuvent retarder considérablement l’évaporation de l’eau. Des études récentes ont montré que de tels films monomoléculaires modifient également la température de la surface de l’eau, la réduction du taux d’évaporation provoquant un réchauffement de la surface. En plus de leur effet sur l’évaporation, les films monomoléculaires modifient également la tension superficielle et la viscosité superficielle d’un substrat aqueux. La présente étude montre que les modifications de ces deux propriétés, outre l’effet de la monocouche sur l’évaporation, peuvent entraîner des modifications de la température de la surface de l’eau.
L’effet des films monomoléculaires sur la température de surface et le mouvement convectif à l’interface eau/air
https://www.sciencedirect.com/science/article/abs/pii/0095852262900193 [10]
Lorsque les conditions sont propices à un taux élevé d’évaporation à la surface d’une eau propre, il y aura un refroidissement rapide, généralement accompagné d’un échange convectif d’eau en vrac plus chaude contre de l’eau de surface plus froide et plus dense. Une monocouche peut modifier la température d’une telle surface en modifiant le taux de cet échange convectif, le changement du flux convectif étant dû à la réduction de la tension superficielle ou au changement de viscosité de la couche d’eau superficielle. Il est donc évident que la température de surface d’une couche d’eau dépend de nombreux facteurs environnementaux et peut être influencée de plusieurs manières par l’ajout d’un film monomoléculaire à l’interface.
Propriétés de type biofilm de la surface de la mer et effets prévus sur les échanges de CO 2
https://www.sciencedirect.com/science/article/abs/pii/S0079661115300616 [12]
Parce que la surface de la mer contrôle diverses interactions entre l’océan et l’atmosphère, elle joue un rôle important dans la biogéochimie marine et la régulation du climat. La surface de la mer est la porte d’échange de gaz, de chaleur et de particules ayant un impact sur le climat. Ainsi, afin de déterminer comment l’océan et l’atmosphère interagissent et réagissent aux changements environnementaux à l’échelle mondiale, la caractérisation et la compréhension de la surface de la mer sont essentielles. La partie supérieure de la colonne d’eau est définie comme la microcouche de surface de la mer et connaît de fortes dynamiques spatiales et temporelles, principalement dues au forçage météorologique. Les zones amorties par les vagues à la surface de la mer sont causées par l’accumulation de matières organiques tensioactives et sont définies comme des nappes. Les nappes naturelles sont fréquemment observées mais leurs propriétés biogéochimiques sont mal connues. Dans la présente étude, nous avons trouvé jusqu’à 40 fois plus de particules d’exopolymère transparentes (TEP), à la base de tout biofilm, dans les nappes par rapport à l’eau sous-jacente. De plus, les nappes étaient caractérisées par une importante biomasse microbienne, une autre caractéristique commune aux biofilms conventionnels sur les surfaces solides. Par rapport aux échantillons ne provenant pas de nappes (similitude moyenne par paire de 70 %), la composition de la communauté de bactéries dans les nappes était de plus en plus différente (similarité moyenne par paire de 45 %) de celle des communautés d’eau en vrac, ce qui indique que la matrice TEP crée des environnements spécifiques pour ses habitants. La distribution potentielle et la fréquence de la formation de nappes dans les régions côtières et océaniques, ainsi que leur effet sur les échanges de CO 2 air-mer , sur la base des données de la littérature. Nous estimons que les nappes peuvent réduire les flux de CO 2 jusqu’à 15 % et jouent donc un rôle local et régional important dans la régulation des interactions air-mer. Au cours des 60 dernières années, l’océan a subi des changements majeurs dus aux émissions anthropiques de gaz à effet de serre, de polluants et d’aérosols. Les changements les plus évidents sont le réchauffement et l’élévation du niveau de la mer, l’acidification des océans et la répartition mondiale des produits chimiques fabriqués par l’homme. Les interactions entre l’océan et l’atmosphère contrôlent l’ampleur et la répartition de ces changements à l’échelle mondiale.
En revanche, les nappes sont des phénomènes de surface de la mer formés par l’action d’amortissement des vagues d’un excès de matière organique tensioactive et scientifiquement définies comme des taches ou des stries lisses et visibles à la surface de la mer. La matière organique peut s’accumuler dans les nappes à un point tel que des plaques de mousse stables peuvent se former. La chimie et la biologie du SML sont souvent mesurables distinctes de l’eau sous-jacente bien mélangée. Grâce à sa position et ses propriétés uniques, le SML est devenu le centre d’une série de processus biogéochimiques et liés au climat, notamment les échanges gazeux air-mer. Cependant, le SML a souvent été négligé dans les efforts visant à comprendre les changements globaux. Par exemple, ses propriétés sont mal caractérisées dans les modèles océaniques et climatiques existants.
Effet de l’accumulation d’hydrocarbures aromatiques polycycliques dans la microcouche de surface de la mer sur leurs échanges air-mer côtiers
https://www.sciencedirect.com/science/article/abs/pii/S0924796309002760 [30]
Parmi la grande variété de substances dangereuses que reçoivent les zones côtières, les hydrocarbures aromatiques polycycliques (HAP) sont reconnus comme une préoccupation majeure en raison de leur accumulation potentielle et de leurs effets mutagènes et cancérigènes sur les organismes. Les apports de HAP peuvent être soit directs (par exemple les rejets d’eaux usées, le ruissellement continental, les marées noires directes et les opérations de transport de pétrole) ou indirects (par exemple les dépôts atmosphériques et les échanges gazeux air-mer), et peuvent subir une série de processus (par exemple dépôt, volatilisation, naufrage). , remise en suspension et dégradation) qui déterminent leur devenir ultime dans l’environnement.
Le transport de contaminants, dont les HAP, entre l’atmosphère et les océans fait l’objet d’une attention particulière depuis le début des années 80 (GESAMP, 1980, GESAMP, 1985). Cependant, les processus physiques et biochimiques contrôlant les échanges air-mer ne sont pas encore bien établis (Upstill-Goddard, 2006). Ces processus peuvent être évalués par modélisation avec un certain degré de précision (Palm et al., 2004), mais des études sur le terrain sont encore nécessaires pour bien comprendre les paramètres clés.
Les paramétrages permettant d’estimer les flux air-mer de contaminants organiques (par exemple PCB et HAP) ont été directement adaptés des études sur les échanges air-mer de CO 2 , mais en raison de leurs différentes propriétés physicochimiques, des données expérimentales sont nécessaires pour élucider les mécanismes impliqués. dans le transport de contaminants organiques. Dans ce cas, l’échange air-mer doit prendre en compte à la fois les processus de transport dissous et associés aux particules, sans négliger d’autres voies, telles que leur accumulation dans la microcouche de surface de la mer (SML), l’effondrement des bulles de gaz ou le transport des aérosols marins vers l’atmosphère ( Cincinelli et al., 2001, Marcomini et al., 2001, Saint-Louis et Pelletier, 2004).
Le rôle du SML en tant que compartiment côtier ne peut être négligé puisqu’il a été reconnu qu’il concentre des contaminants hydrophobes et d’autres substances et affecte les échanges air-mer par l’apparition de films organiques biogéniques et anthropiques. Ces films sont variables dans le temps et dans l’espace, ce qui rend difficile une compréhension globale du rôle du compartiment SML dans le transport global des contaminants. De plus, le SML a été impliqué dans la modification des flux de gaz biogènes tels que le méthane à travers l’interface air-mer, probablement en raison de l’activité microbienne dans le SML.
La cultivabilité bactérienne dans les habitats marins oligotrophes à mésotrophes varie généralement de 0,001 % à 1 %, alors que ce pourcentage peut augmenter dans les environnements riches en nutriments (par exemple, boues activées).
Prédire la bioaccumulation de composés organiques dans les organismes marins à l’aide des coefficients de partage octanol/eau
https://www.sciencedirect.com/science/article/abs/pii/0025326X83906045 [31]
Les sédiments et les animaux collectés à proximité de la zone de rejet de la station d’épuration des eaux usées du comté de Los Angeles ont été analysés pour détecter 27 composés organiques sélectionnés qui avaient été identifiés dans les effluents. Il a été constaté que les concentrations de ces composés organiques dans les sédiments et les tissus étaient positivement corrélées entre elles et avec le coefficient de partage n -octanol/eau, mais qu’elles étaient négativement corrélées avec la concentration de l’effluent. Il a été conclu que les coefficients de partage n -octanol/eau devraient être utilisés pour prédire quels composés organiques ont le potentiel de se bioaccumuler et donc de provoquer des effets biologiques.
Tensioactifs synthétiques en milieu marin
https://www.sciencedirect.com/science/article/abs/pii/B9780323902977000093 [18]
Les tensioactifs synthétiques font partie des produits chimiques produits et consommés en plus grandes quantités dans le monde, en raison de leur variété d’applications, principalement comme ingrédients clés dans les détergents et les nettoyants, mais aussi comme agents mouillants, additifs pour peinture et désinfectants, entre autres utilisations. . Ce chapitre donne un aperçu des techniques analytiques utilisées pour leur détermination dans des échantillons marins solides et liquides, ainsi que des concentrations trouvées dans différents écosystèmes marins. Le devenir des tensioactifs est également discuté en termes de sorption, de dégradation et de bioaccumulation. Les principaux aspects du potentiel toxique des tensioactifs, y compris les effets aigus et chroniques, sont également présentés.
Fuite de tensioactifs dans les eaux grises : impact environnemental
https://www.sciencedirect.com/science/article/abs/pii/S2213343723014549 [17]
Les eaux grises se caractérisent par la présence de matières organiques, de savons, d’huiles, de nutriments, de micro-organismes, de matières solides en suspension, de tensioactifs et de minéraux. Les tensioactifs constituent un facteur de risque important parmi la diversité des contaminants. Ces tensioactifs proviennent de diverses sources telles que les textiles, les peintures, les produits de beauté, les produits pharmaceutiques et l’industrie du papier. Les tensioactifs sont des molécules composées de parties hydrophiles et hydrophobes et sont généralement destinées à être utilisées comme agent de nettoyage dans les détergents et les savons. Les tensioactifs sont généralement de trois types cationiques, anioniques et non ioniques. Les tensioactifs tels que les LAS, les éthoxylates d’alkylphénol (APE), les alkyléthoxysulfates (AES) et les composés d’ammonium quaternaire sont généralement présents dans les effluents d’eaux grises. La libération de ces tensioactifs constitue une double menace, à la fois en termes d’implications environnementales et de risques potentiels pour les organismes de l’écosystème. Cette revue consolide plusieurs aspects relatifs aux tensioactifs provenant des effluents de GW. Il offre un aperçu fondamental de la composition des eaux grises, les tensioactifs étant identifiés comme présentant le niveau de risque le plus élevé. L’examen souligne également l’importance d’atténuer les fuites de tensioactifs des eaux usées dans l’environnement, car elles ont un impact sur la qualité des sols, les écosystèmes aquatiques et divers organismes. Les tensioactifs sont fréquemment présents dans les produits d’entretien ménager et les articles de toilette. La tension superficielle entre deux liquides ou entre un liquide et un solide peut être réduite par ces tensioactifs. Les tensioactifs contenus dans GW peuvent affecter négativement les caractéristiques du sol et la croissance des plantes. Les GW contenant des tensioactifs, lorsqu’ils sont rejetés dans le sol, peuvent réduire sa capacité à retenir l’humidité et les minéraux. En effet, les agrégats du sol, qui sont cruciaux pour préserver une structure et une porosité saines du sol, peuvent être brisés. Les tensioactifs sont les principaux contaminants des GW avec des propriétés diverses. Ces tensioactifs peuvent avoir un double impact en fonction de leur type, de leur concentration et d’autres paramètres opérationnels. En règle générale, les eaux usées hautement concentrées peuvent avoir des effets néfastes sur l’environnement et les organismes vivants. Des techniques conventionnelles et avancées doivent donc être explorées pour atteindre une efficacité maximale. Cependant, l’élimination complète des tensioactifs reste un défi en raison de problèmes tels qu’une faible efficacité des systèmes de traitement.
Composition en matière organique et enrichissement en polluants de la microcouche de surface de la mer à l’intérieur et à l’extérieur des nappes
https://www.sciencedirect.com/science/article/abs/pii/014111369390100E [13]
Une relation étroite apparaît entre l’accumulation de polluants et la baisse de la tension superficielle des nappes. Cela a une conséquence directe : l’effet amortisseur des vagues qui lisse la surface de la mer, rendant les nappes observables, caractéristiques de l’accumulation de polluants.
Métabolites persistants des polyéthoxylates d’alkylphénol dans le milieu marin
https://www.sciencedirect.com/science/article/abs/pii/030442039090020D
Les métabolites persistants des polyéthoxylates de nonylphénol (NP n EO, n = 1–18), à savoir le nonylphénol (NP), le monoéthoxylate de nonylphénol (NP1EO) et le diéthoxylate de nonylphénol (NP2EO), ont été analysés dans des échantillons marins, notamment des sédiments et des sédiments artificiellement remis en suspension. et de l’eau. La détermination par chromatographie liquide haute performance par fluorescence UV a été réalisée après extraction au Soxhlet à l’hexane des matrices sédimentaires suivie d’un nettoyage sur minicolonnes d’aminosilice. Quatre campagnes d’échantillonnage dans cinq stations représentatives de la lagune de Venise ont été menées pour vérifier la variabilité temporelle et spatiale des produits chimiques examinés. Un dispositif de remise en suspension portable nous a permis d’analyser la première couche de sédiments de 0,01 à 0,15 mm, où la somme de NP, NP1EO et NP2EO était comprise entre 0,15 et 13,7 μg g −1 (en poids sec), soit au moins cinq fois supérieure à dans les 5 cm de sédiments sous-jacents. Les quantités de matières remises en suspension et les concentrations de NP, NP1EO, NP2EO qui y sont liées ont montré une dépendance saisonnière marquée : jusqu’à deux fois la matière remise en suspension, par unité de surface de sédiment, a été mesurée en avril et juillet, par rapport à celle de février, mais les NP, NP1EO et NP2EO remis en suspension par unité de surface de sédiment étaient en février d’un ordre de grandeur plus élevé. Un facteur majeur responsable de cette tendance était lié à la prolifération de macroalgues qui contenaient une concentration moyenne de NP+NP1EO+NP2EO de 0,25 ± 0,15 μg g −1 (poids sec). Dans l’eau, des oligomères NPEO comportant jusqu’à 13 unités éthoxy ont été trouvés dans une plage de concentrations globale de 0,6 à 4,5 μg l −1 .
Risques environnementaux et toxicité des tensioactifs : aperçu des techniques d’analyse, d’évaluation et de remédiation
https://link.springer.com/article/10.1007/s11356-021-16483-w [ ]
Les tensioactifs font partie des contaminants émergents les plus problématiques qui sont continuellement rejetés dans l’environnement par les usines de traitement des eaux usées (STEP).
Ce travail a examiné de manière approfondie la toxicité et les risques de divers tensioactifs et de leurs produits dégradés dans les matrices environnementales, diverses procédures analytiques et méthodes d’assainissement de ces tensioactifs. Les résultats ont révélé que la concentration élevée de tensioactifs et de leurs produits dégradés perturbe la dynamique microbienne et leurs processus biogéochimiques importants, entrave les processus de survie des plantes et leur niche écologique, et retarde les fonctionnalités organiques et systémiques humaines. Les énormes effets néfastes des tensioactifs sur la santé et l’environnement nécessitent de développer, sélectionner et faire progresser les diverses techniques d’analyse et d’évaluation pour parvenir à une identification et une quantification efficaces de plusieurs tensioactifs dans différentes matrices environnementales.
Cependant, la persistance des produits transformés dans un environnement est très préoccupante pour la durabilité environnementale et la santé des écosystèmes.
En raison principalement des applications étendues des tensioactifs, leur énorme concentration provenant principalement des eaux usées urbaines ou industrielles et domestiques peut aboutir dans les stations d’épuration municipales (STEP) ou être directement rejetée dans l’environnement. Il y a plusieurs années, on estimait que les tensioactifs présents dans les eaux usées domestiques et industrielles se situaient respectivement entre 1 et 10 mg/L et 300 mg/L. Il est possible que les tensioactifs pénètrent dans l’eau potable par les stations d’épuration des eaux usées, posant ainsi un risque pour la santé humaine, animale et aquatique. De plus, le devenir et la structure chimique des surfactants, des systèmes environnementaux et des accepteurs d’électrons contribuent principalement à la persistance, à la dégradabilité et aux effets écologiques des surfactants dans l’environnement.
Certains tensioactifs disponibles dans le commerce constituent une grave menace environnementale et publique pour les humains et les écosystèmes. Par exemple, les tensioactifs anioniques, principalement les LAS, provoquent des impacts biochimiques, pathologiques, physiologiques et autres sur les écosystèmes aquatiques/terrestres. De plus, le LAS provoque des irritations cutanées et des problèmes respiratoires et réduit la résistance du biote aquatique au stress environnemental, à la reproduction et aux processus de croissance. Les tensioactifs augmentent généralement la solubilité des contaminants et facilitent ainsi l’eutrophisation. De plus, les propriétés hydrophobes croissantes des tensioactifs augmentent proportionnellement leur toxicité. Ces vastes impacts suscitent par conséquent des inquiétudes en matière de santé publique et d’environnement concernant la forte concentration de tensioactifs.
Des études antérieures ont montré qu’une forte concentration de tensioactifs et leurs produits dégradés se déposent dans divers compartiments environnementaux en raison de leurs applications étendues. Les tensioactifs peuvent pénétrer dans l’environnement à partir des effluents générés par les produits agrochimiques, les produits industriels et les activités domestiques et par conséquent provoquer une pollution environnementale. Par exemple, les biocides, les herbicides et les pesticides font partie des produits agrochimiques, tandis que les produits de soins personnels, les émulsifiants, les agents mouillants, les détergents et les revêtements ou adoucissants des tissus, du papier et des tapis sont des produits industriels notables qui contribuent continuellement à la protection de l’environnement grâce aux tensioactifs. pollution. De plus, la lessive, la désinfection et la fumigation sont d’importantes activités domestiques qui rejettent des tensioactifs dans l’environnement.
Le devenir, la distribution et la persistance des tensioactifs dans l’environnement sont principalement influencés par la sorption et la bio/photodégradation. Ces processus dépendent principalement de divers facteurs environnementaux tels que le pH, la température et la salinité.
Plusieurs études ont montré de graves impacts écologiques et sanitaires des tensioactifs toxiques sur les humains, d’autres vertébrés, la faune du sol, les micro-organismes, les crustacés et les plantes terrestres. Par exemple, la formation de mousse et la sédimentation des tensioactifs détériorent la qualité de l’eau en réduisant le transfert d’oxygène de l’air ou de l’eau et la capacité autonettoyante des rivières, à des degrés différents selon les classes de tensioactifs. De plus, les tensioactifs améliorent la solubilité des polluants organiques persistants (POP) dans la phase aqueuse, et les produits d’aérosols et de tensioactifs qui en résultent ont un impact significatif sur l’atmosphère et le climat. Les changements dans les activités physiologiques et biochimiques des organismes aquatiques par le LAS retardent leur métabolisme et leur croissance, endommagent la membrane cellulaire et provoquent la rupture du complexe protéique de la chlorophylle.
La concentration médiane efficace (EC50) et la concentration médiane d’inhibition de la croissance (IC50) déterminent respectivement le risque et la toxicité des tensioactifs sur les animaux, les plantes et les micro-organismes. Par exemple, Ge et al. ( 2010 ) ont découvert que la CE50 à 96 h du chlorure d’acétyltriméthylammonium sur la microalgue verte Chlorella vulgaris est de 145 ± 13,35 μg/L, tandis que la CE50 du benzalkonium sur les invertébrés est de 5,90 μg/L. De plus, la CI50 du chlorure de benzyldiméthyldodécylammonium pour les bactéries est de 170,0 μg/L. Les tensioactifs peuvent retarder la croissance microbienne et augmenter les mutations microbiennes et la mortalité. Par exemple, les éthoxylates de nonylphénol (NPEO) peuvent dissocier la production d’énergie, ce qui peut par conséquent perturber les processus de croissance microbienne et de nitrification. De plus, les tensioactifs anioniques perturbent les structures internes et les fonctions microbiennes telles que la résistance environnementale, la croissance, le stress compétitif et la reproduction. L’absorption de tensioactifs par un micro-organisme peut dépolariser la membrane cellulaire microbienne et diminuer l’absorption des nutriments, l’acceptation de l’oxygène et la libération des métabolites toxiques. Certains tensioactifs ont de graves conséquences sur la santé humaine en raison de l’ingestion ou de la consommation d’aliments contaminés. Par exemple, les tensioactifs réagissent avec les protéines existantes dans le foie et le sérum, provoquant ainsi des effets métaboliques à long terme et une perturbation du système endocrinien humain. De même, il a été rapporté que certains tensioactifs provoquent des brûlures ou des irritations de la peau humaine ainsi que des problèmes oculaires et respiratoires. Les éthoxylates et carboxylates d’alkylphénol retardent la pénétration souterraine des composés pharmaceutiques. Les éthoxylates de nonylphénol ont des effets œstrogéniques sur les amphibiens, les mammifères et les Poissons. Certains tensioactifs tels que le LAS endommagent la membrane des cellules racinaires et modifient la structure de la membrane. Par conséquent, cela a altéré la transpiration et la translocation des nutriments essentiels et de l’eau.
En raison des risques que représentent les tensioactifs et leurs produits dégradés pour l’environnement, il est pertinent de déterminer leurs concentrations dans les matrices environnementales. Cependant, l’analyse en laboratoire des tensioactifs pourrait s’avérer une tâche difficile à réaliser. La raison en est que les tensioactifs se trouvent généralement dans des mélanges comprenant différents composants tels que des isomères, des homologues et d’autres impuretés. De plus, les tensioactifs sont souvent présents en quantités infimes, généralement inférieures aux limites détectables des instruments d’analyse. La complexité et la variété des matrices dans lesquelles les tensioactifs sont naturellement présents compliquent et entravent en outre leur analyse. À cette fin, une technique sophistiquée de pointe est nécessaire pour effectuer l’analyse des tensioactifs dans des flux aqueux.
La présence de tensioactifs dans les environnements aquatiques et terrestres a des effets néfastes sur les micro-organismes, les plantes, les animaux et les humains. L’immense utilisation des tensioactifs, leurs risques pour la santé et les politiques réglementaires strictes concernant leur limite autorisée nécessitent leur contrôle dans l’environnement. Cette limite admissible a fait l’objet de débats entre plusieurs chercheurs. Par conséquent, plusieurs approches d’assainissement ont été recommandées pour nettoyer les tensioactifs présents dans l’environnement. Au cours des dernières décennies, la bioremédiation a souvent été utilisée pour éliminer les tensioactifs de la matrice environnementale. Le sonoréacteur pour l’assainissement des tensioactifs des eaux usées présente de nombreux avantages : mode de fonctionnement simple, plus respectueux de l’environnement, utilisation minimale de produits chimiques, absence de génération de boues et de sous-produits. De plus, le faible coût d’exploitation et de maintenance en fait un processus de traitement d’oxydation avancé et efficace. En principe, ce réacteur fonctionne sur le principe de la génération de radicaux libres. Récemment, Hadi et al. (Dehghani et al. 2019 ) ont appliqué un sonoreactor pour dégrader le LAS à partir d’une solution aqueuse. Ils ont constaté que la dégradation du LAS augmente avec le temps. À l’inverse, une augmentation de la concentration initiale de LAS diminue le taux d’élimination du LAS. En revanche, l’augmentation de la puissance acoustique entraîne une dégradation plus importante du LAS. Ceci est dû à la réaction possible dans la cavitation de l’acteur sonore. Cependant, une puissance acoustique élevée impliquait une énergie plus élevée. Il convient donc de maintenir un équilibre entre efficacité et coût énergétique.
Concentrations de contaminants et toxicité de la microcouche de surface de la mer près de Los Angeles
https://www.sciencedirect.com/science/article/abs/pii/0141113687900249 [19]
Des échantillons de microcouches de surface de la mer ont été collectés dans six zones côtières recevant différentes quantités d’apports anthropiques. Les échantillons ont été analysés pour détecter certains métaux traces, hydrocarbures chlorés et hydrocarbures aromatiques polycycliques. Les toxicités relatives des échantillons ont été déterminées à l’aide d’essais biologiques sur des embryons de poisson.
Les concentrations de contaminants ont généralement augmenté des stations offshore vers les stations côtières. Les concentrations de contaminants étaient plusieurs fois plus élevées dans les échantillons de microcouches provenant des ports hautement industrialisés de Los Angeles et de Long Beach que dans les échantillons provenant d’un site situé à 15 km au large. Les échantillons de microcouches provenant des stations côtières étaient significativement plus toxiques et induisaient beaucoup plus d’anomalies de développement et d’aberrations chromosomiques que les échantillons provenant des stations offshore.
Réponse des particules submitochondriales aux sulfonates d’alkylbenzène linéaires, aux polyéthoxylates de nonylphénol et à leurs dérivés de biodégradation
https://setac.onlinelibrary.wiley.com/doi/10.1002/etc.5620130507 [32]
Les effets sur les paramètres respiratoires mitochondriaux des LAS, des polyéthoxylates de nonylphénol (NPEO) et de certains de leurs produits de biotransformation, à savoir les carboxylates de sulfophényle (SPC), le nonylphénol (NP) et l’acide nonylphénoxy acétique (NP1EC), ont été enregistré en utilisant la réponse in vitro des particules soumetchondriales (SMP) du cœur de bœuf. La toxicité de ces composés a été estimée en déterminant leurs effets sur le transfert d’électrons inverse couplé à l’énergie (RET), qui est induit par l’ATP et le succinate au premier niveau de la chaîne respiratoire et réduit le NAD + exogène en NADH. La toxicité des substances, exprimée en concentration de toxique diminuant le taux de réduction du NAD + jusqu’à 50 % (CE50), variait de 0,61 mg/L pour un mélange LAS commercial à 18 000 mg/L pour des RCP individuels ; de 1,3 mg/L pour NPEO, avec une moyenne de 10 unités éthoxy, à 8,2 et 1,8 mg/L pour NP1EC et NP, respectivement. Ces résultats ont été liés à la structure moléculaire de chaque classe de composés et comparés aux valeurs de toxicité obtenues par divers systèmes biologiques actuellement utilisés pour les tests de toxicité. Les données sur la toxicité aiguë ont démontré que (a) le test biologique SMP permet de reproduire la réponse toxicologique d’organismes entiers, tels que les poissons et les invertébrés, aux produits chimiques testés ; et (b) la partie hydrophobe de ces composés joue un rôle important dans le déclenchement de leurs effets toxiques. D’un point de vue toxicologique, il faut prêter attention à la présence dans les eaux naturelles de LAS résiduels, alors que dans le cas du NPEO, tant le tensioactif non altéré que tous les produits de biotransformation doivent être identifiés et quantifiés.
Distribution et devenir diagénétique des tensioactifs synthétiques et de leurs métabolites dans les sédiments estuariens contaminés par les eaux usées
https://www.sciencedirect.com/science/article/abs/pii/S0269749118316658?via%3Dihub [33]
Les tensioactifs sont des produits chimiques produits en grande quantité et utilisés dans de nombreuses applications domestiques et industrielles et, après utilisation, ils sont les plus abondants organiques contaminants dans les eaux usées. Leur décharge pourrait mettre en danger le récepteur aquatique ecosystems, y compris les sédiments, où ils ont tendance à s’accumuler. Il s’agit de la première étude complète sur leur distribution et leur devenir dans ce compartiment environnemental puisque nous avons effectué une analyse simultanée des trois principales classes de tensioactifs (anioniques :LAS; non ionique : NPEO et AEO ; cationique : DTDMAC, BAC et ATMAC et certains de leurs transformation products(SPC, NP, NPEC et PEG). Pour tenir compte des tendances spatiales et temporelles, des sédiments de surface et des carottes datées ont été collectés dans la baie de Jamaica, un estuaire fortement pollué par les eaux usées de la ville de New York. Les concentrations de tensioactifs dans les sédiments de surface étaient comprises entre 18 et > 200 μg g −1 et présentaient une légère variation (<10 %) au cours des différentes années d’échantillonnage (1998, 2003 et 2008). Les tensioactifs cationiques ont été trouvés aux concentrations les plus élevées, le DTDMAC représentant entre 52 et 90 % de la somme totale des composés cibles. Profils de concentration verticale dans des carottes datées de la station la plus contaminée, à proximité du plus grand local sewage treatment(STP), a indiqué deux pics de tensioactifs souterrains au milieu des années 1960 (469 μg g−1) et fin des années 1980 (572 μg g−1) coïncidant avec les mises à niveau STP connues. Cette tendance a été observée pour la plupart des composés cibles, à l’exception du DADMAC , du C22ATMAC et du PEG, qui ont montré une augmentation continue vers le haut des noyaux. La dégradation in situ a été étudiée en comparant sediment core des échantillons prélevés à 12 ans d’intervalle (1996 et 2008) ont révélé une diminution nette du PEG et des tensioactifs spécifiques (BAC, ATMAC, NPEO et AEO) accompagnée de concentrations croissantes de métabolites (SPC, NP et NPEC). DTDMAC, DADMAC et LAS sont cependant restés stables au cours de cette période, suggérant un comportement récalcitrant sous le régime anaerobic conditions dans les sédiments de Jamaica Bay.
En raison de leur utilisation intensive, l’analyse des eaux usées indique que les tensioactifs sont les substances organiques prédominantes en termes de concentration et de fréquence . Bien que la plupart des tensioactifs soient efficacement éliminés dans les stations d’épuration des eaux usées (STP), des concentrations de l’ordre de quelques microgrammes par litre peuvent être trouvées dans les eaux de surface réceptrices. Leurs niveaux augmentent dans les sédiments car ils constituent des puits naturels pour les tensioactifs en raison de l’affinité relativement élevée de ces produits chimiques pour les particules de carbone organique et/ou les surfaces chargées. En raison de leur utilisation répandue, de la spécificité de leur source et de leur taux de dégradation généralement faible dans des conditions anaérobies, de nombreux tensioactifs peuvent être utilisés comme indicateurs moléculaires dans les sédiments pour surveiller la contamination causée par les activités humaines. Les comportements environnementaux des tensioactifs, par exemple la biodégradation et les interactions avec d’autres contaminants organiques, ont été bien étudiés dans des recherches antérieures, cependant, le comportement à long terme de ces composés dans ce compartiment environnemental est encore largement inconnu. mais essentiel pour comprendre leur sort final dans les systèmes aquatiques.
Bien qu’ayant des volumes de production inférieurs à ceux de leurs homologues anioniques et non ioniques, l’analyse des tensioactifs cationiques tels que les composés d’ammonium quaternaire (QAC) a révélé des niveaux plus élevés dans les sédiments. Certains d’entre eux (par exemple, le chlorure d’alkyltriméthyle et de ditallowdiméthylammonium, ou ATMAC et DTDMAC, respectivement) ont été identifiés comme contaminants émergents dans les boues d’épuration et les sédiments estuariens, atteignant des concentrations allant jusqu’à des milliers de microgrammes par gramme de sédiment. Ces concentrations observées sont si élevées principalement en raison de la combinaison d’une forte capacité d’adsorption résultant de leur nature hydrophobe et cationique et d’une dégradation microbienne minime en l’absence d’oxygène. Une caractérisation très limitée est disponible pour d’autres QAC tels que les chlorures de benzylalkyldiméthylammonium (BAC) et les chlorures de dialkyldiméthylammonium (DADMAC), avec des longueurs de chaîne n-alkyle plus courtes variant entre C8 et C10. Les deux sont utilisés dans les désinfectants et leurs impacts potentiels sur les communautés microbiennes et sur la propagation des gènes de résistance au QAC sont inconnus. En outre, de nouveaux types de tensioactifs tels que les amines de suif polyoxyéthylène, le sulfate d’éther de diglycol et le chlorure de béhentrimonium ont été signalés dans l’environnement ces dernières années en raison de l’augmentation production et utilisation.
Dégradation incomplète des tensioactifs linéaires alkylbenzène sulfonates dans les eaux de surface brésiliennes et poursuite de leurs métabolites polaires dans les eaux potables
https://www.sciencedirect.com/science/article/abs/pii/S0048969701008737
Au Brésil, plus de 90 % de la population n’est pas raccordée aux stations d’épuration municipales . En conséquence, les eaux de surface reçoivent en permanence des quantités considérables d’eaux usées domestiques non traitées contenant des tensioactifs comme constituant majeur. Ces eaux polluées suscitent un intérêt particulier si elles sont utilisées comme source de production d’eau potable. Dans ce travail, la rivière Rio Macacu a été surveillée pour détecter la présence du tensioactif anionique linéaire alkylbenzène sulfonate (LAS) le plus largement utilisé, ainsi que de son principal produit de dégradation, les sulfophénylcarboxylates (SPC). Afin de suivre le devenir des deux composés après leur émission dans la rivière, des échantillons ont été collectés à plusieurs endroits le long de la berge et analysés par chromatographie liquide-spectrométrie de masse après enrichissement par extraction en phase solide. Les concentrations de LAS variaient entre 14 et 155 μg l −1 et les niveaux de leurs intermédiaires métaboliques étaient compris entre 1,2 et 14 μg l −1 . La capacité d’auto-épuration de l’eau a été démontrée de manière impressionnante dans le cours supérieur de la rivière en aval d’une ville considérée comme un point de rejet majeur, tandis que dans le cours inférieur, des concentrations relativement constantes des deux analytes ont été détectées, ce qui s’explique par une augmentation globale niveau de pollution. En outre, une série d’échantillons d’eau potable de Niterói et São Gonçalo, fournis par les mêmes usines de traitement des eaux de surface du Rio Macacu, ont été prélevés au cours de deux périodes d’échantillonnage et examinés pour détecter la présence du SPC fortement polaire, soupçonné de contourner les processus de purification. Les niveaux détectés dans l’eau potable variaient entre 1,6 et 3,3 μg l −1 . Pour les analyses des eaux potables et des eaux de surface, la configuration des pics d’un homologue sélectionné du SPC composé de plusieurs isomères de position a servi d’indicateur pour décrire la progression de la dégradation du SPC survenue dans la rivière et a pu être utilisée pour distinguer les eaux potables de différentes origines.
Les alkylbenzènesulfonates linéaires (LAS) sont les tensioactifs anioniques les plus importants largement utilisés dans la formulation de détergents ménagers et de produits de nettoyage industriels et institutionnels. La quasi-totalité des quelque 2,4 millions de tonnes consommées chaque année dans le monde est éliminée après utilisation dans les eaux usées qui peuvent être traitées dans les stations d’épuration municipales. De nombreuses études menées sur le devenir des LAS lors du traitement des eaux usées ont indiqué qu’ils sont efficacement éliminés par des processus physiques, chimiques et biologiques. Outre la précipitation et l’adsorption sur les matières en suspension, qui peuvent varier de 30 à 70 %, la dégradation microbienne représente généralement la principale voie d’élimination, généralement environ 80 %, ce qui entraîne une réduction globale de la charge de LAS de 95 à 99,5 % dans les systèmes à boues activées. Néanmoins, certains résidus du tensioactif intact ainsi que ses intermédiaires de dégradation aérobie, les sulfophénylcarboxylates (SPC), pénètrent dans les eaux réceptrices via les sorties de la STEP. Compte tenu des énormes quantités de LAS utilisées, les concentrations dans les eaux de surface se situent dans la plage inférieure de μg l −1.
En revanche, si les eaux usées domestiques sont déversées directement dans les cours d’eau naturels en raison d’installations de traitement déficientes, les niveaux peuvent être considérablement plus élevés. Alors qu’en Europe occidentale et aux États-Unis, la majorité des ménages sont raccordés à des stations d’épuration, le rejet d’eaux usées non traitées dans les rivières est encore largement pratiqué dans de nombreux pays. Cela est particulièrement préoccupant puisque dans ces circonstances, les organismes aquatiques sont exposés à des niveaux considérables de tensioactifs, qui présentent des toxicités relativement élevées. Quant au devenir des tensioactifs après rejet d’eaux usées non traitées, leur concentration dans l’eau pourrait être réduite par sorption sur les sédiments fluviaux, ainsi que par biodégradation par les communautés bactériennes endogènes présentes dans le cours d’eau – avec une évolution plus lente. cinétique par rapport à la STEP mais la forte solubilité dans l’eau des LAS et de leurs métabolites encore plus polaires permettent leur transport par convection sur des distances relativement longues. En fin de compte, l’embouchure des rivières polluées dans les estuaires puis dans la mer contribue à la contamination des eaux côtières. La grande mobilité des composés aromatiques sulfonés les rend également potentiellement dangereux en termes de contamination des ressources en eaux souterraines et suscite des inquiétudes quant à la qualité de l’eau potable.
Concernant la détermination du LAS et du SPC dans les matrices environnementales, la complexité du mélange de tensioactifs commerciaux, composé de quatre homologues alkyles, chacun d’eux comprenant plusieurs isomères phényles ainsi que les intermédiaires de dégradation nécessite l’application de méthodes analytiques sophistiquées.
Le rôle des tensioactifs dans le traitement des eaux usées : impact, élimination et techniques futures : une revue critique
https://www.sciencedirect.com/science/article/abs/pii/S0043135418307541?via%3Dihub [34]
La responsabilité importante de réagir à l’évolution des déchets produits par les consommateurs et par l’industrie qui posent des défis environnementaux. Les tensioactifs en font partie des contaminants, Ils sont intéressants en raison de leur utilisation domestique et industrielle de plus en plus omniprésente et de la difficulté que leur présence entraîne pour le traitement traditionnel. En réponse à ce domaine en développement, cette revue critique prend en compte la recherche dans divers domaines techniques pour fournir un aperçu actualisé de l’impact des tensioactifs sur l’environnement, la santé et leur élimination. Cela a révélé que les préoccupations majeures concernant les tensioactifs sur l’environnement et sur la santé ont été corroborées au cours des cinq dernières années. Les recherches actuelles sur l’élimination se concentrent sur l’optimisation des traitements biologiques et chimiques existants des eaux usées. Malgré les améliorations apportées aux méthodes biologiques traditionnelles utilisant des prétraitements chimiques, il existe un manque évident de consensus quant à la stratégie idéale. Inconvénients et solutions potentielles pour une série de ces problèmes technologiques, y compris Fenton réaction et la dégradation aérobie, sont discutés. Dans ce domaine, les auteurs recommandent une diversité améliorée des tensioactifs utilisés pour la recherche et comblent d’importantes lacunes dans les connaissances. Les utilisations potentielles des tensioactifs pour optimiser le traitement des eaux usées, telles que les zéolites modifiées par des tensioactifs (SMZ), sont également discutées. Cette revue révèle que l’élimination des tensioactifs des eaux usées est un domaine prometteur et difficile qui mérite des recherches plus approfondies.
Le traitement des contaminants émergents, tels que les tensioactifs, constitue l’un des défis majeurs du traitement des eaux usées. Les tensioactifs constituent un membre particulièrement complexe de ce groupe. Cela est dû à la large utilisation et à la gamme de tensioactifs et à leur impact négatif à la fois sur le processus de traitement des eaux usées et, plus important encore, sur l’environnement.
Les tensioactifs sont des molécules amphiphiles contenant deux parties distinctes : un groupe hydrophile et un groupe lipophile et peuvent être définis au sens large comme des composés qui modifient les relations énergétiques aux interfaces, souvent en termes de modification soit la tension superficielle ou interfaciale. Lorsqu’ils sont présents en faibles concentrations, les tensioactifs réduisent la quantité d’énergie libre interfaciale nécessaire pour modifier l’interface ou la surface et favorisent l’expansion de l’une des interfaces au sein du système, améliorant ainsi l’efficacité de nombreux processus. En conséquence, les tensioactifs sont largement utilisés comme agents mouillants, détergents et agents émulsifiants, tandis que d’autres tensioactifs, tels que les composés d’ammonium quaternaire (QAC), endommagent également la membrane externe des bactéries à Gram négatif et peuvent donc être utilisés comme désinfectants. La structure de base d’un tensioactif.
L’utilisation de tensioactifs est extrêmement courante et a augmenté depuis les années 1980, passant de 1,7 million de tonnes utilisées en 1984 et 9,3 millions de tonnes utilisées en 1995 aux 15,93 millions de tonnes utilisées en 2014 et aux 24,19 millions de tonnes attendues. utilisé en 2022. De nombreuses industries utilisent des tensioactifs, notamment l’industrie textile mais aussi dans les industries pharmaceutique, cosmétique, minière, de récupération du pétrole et du papier même s’il faut noter qu’en 2014, les soins à domicile représentaient pour plus de 40 % du marché mondial.
Cependant, alors que le marché mondial des tensioactifs est en croissance, on s’inquiète également de plus en plus des dommages causés par les tensioactifs à l’environnement, en particulier à cause des eaux usées. De nombreux articles ont été publiés concernant les effets négatifs des tensioactifs sur l’environnement, la faune et les humains. Venhuis et Mehrvar (2004) signalent que les alkylbenzènesulfonates linéaires (LAS) peuvent, entre autres effets, endommager les branchies des poissons, modifier les habitudes de nage des larves de moules bleues et provoquer une sécrétion excessive de mucus ainsi que (chez le gobie commun) réduire la respiration. De nombreuses autres études et revues ont fait état de la toxicité de divers tensioactifs différents ainsi que leurs effets sur l’environnement, principalement en mettant l’accent sur une variété d’organismes aquatiques. Les tests de toxicité aiguë montrent qu’il existe une large gamme de valeurs CL 50 d’une espèce à l’autre et peuvent aller de 210 à 500 000 μg/L pour les tensioactifs non ioniques, de 80 à 2 800 000 μg/L pour les tensioactifs cationiques et de 110 à 92 000 μg/L. pour les tensioactifs anioniques. De nombreux tensioactifs testés se sont révélés suffisamment toxiques pour pouvoir être classés comme très toxiques, toxiques et nocifs selon la directive 67/548/CEE de l’Union européenne.
D’autres études ont noté que les tensioactifs peuvent augmenter la quantité de composés d’hydrocarbures organiques solubilisés dans les sources d’eau ou être absorbés dans les sols et les sédiments où ils pourraient potentiellement devenir nocifs pour les cultures tout en affectant également la dégradation des composés organiques hydrophobes (Ying, 2006). . D’autres études ont montré que, même s’il peut y avoir une élimination importante de certains tensioactifs (plus de 99 % des LAS, le tensioactif restant qui finit dans les rivières et autres sources d’eau peut se bioaccumuler au sein de certaines espèces entraînant une biotransformation des tissus. En outre, il a été démontré que l’exposition à certains tensioactifs, tels que les chlorures de benzalkonium (BAC), peut augmenter la résistance des microbes aux antibiotiques.
Ces préoccupations ont conduit, en Europe, à une réglementation beaucoup plus stricte en termes de biodégradabilité des tensioactifs et de leur teneur en composés phosphorés et en phosphates. Aux États-Unis, tous les tensioactifs qui contournent le traitement des eaux usées doivent répondre à des critères spécifiques. Malgré le renforcement de la réglementation, l’élimination des tensioactifs des eaux usées reste une préoccupation croissante, surtout compte tenu de la forte toxicité et de l’omniprésence des tensioactifs. Cela se reflète dans l’augmentation constante du nombre de publications traitant à la fois des tensioactifs et du traitement des eaux usées.
Cependant, il convient de noter au lecteur que bon nombre de ces publications ne considèrent pas les tensioactifs comme leur objectif principal. En général, on constate un manque d’attention portée aux problèmes posés par les tensioactifs dans le processus de traitement des eaux usées, malgré l’augmentation globale des publications observées dans la littérature. Un aperçu global de l’impact des tensioactifs sur le processus de traitement des eaux usées est souvent absent de la littérature traitant du sujet.
Il est donc nécessaire de procéder à une révision actualisée du sujet. Cette revue examinera l’étendue des méthodes de traitement actuelles utilisées pour éliminer les tensioactifs anioniques, cationiques et non ioniques couramment utilisés, les travaux dans la littérature pour améliorer ces techniques et les nouvelles tentatives et stratégies d’élimination pour utiliser les tensioactifs dans le traitement des eaux usées. L’objectif est que cette approche motive des recherches plus approfondies dans les domaines prometteurs découverts dans cette revue pour aborder cet important problème émergent dans l’ingénierie des eaux usées.
Effet de l’inhibition des tensioactifs anioniques sur le traitement des eaux usées par un bioréacteur à membrane anaérobie immergée : efficacité, activité des boues et récupération du méthane
https://www.sciencedirect.com/science/article/abs/pii/S1385894717300165 [35]
À l’heure actuelle, les procédés conventionnels de traitement des eaux usées, tels que les techniques aérobies et anoxiques, ne sont pas durables en raison de leur grande consommation d’énergie, de leurs grandes qualités de production de résidus et de leur incapacité à récupérer les ressources potentielles disponibles dans les eaux usées. Le bioréacteur à membrane anaérobie immergé (SAnMBR) offre une stratégie alternative pour le traitement des eaux usées en raison de sa rétention presque absolue de biomasse, de son potentiel à générer un effluent de meilleure qualité, de sa consommation d’énergie réduite et de sa génération de biogaz. Il a été étudié avec succès pour le traitement des eaux usées à haute concentration, telles que les eaux usées de l’industrie alimentaire et des boissons et le fumier de porc. Ce n’est que récemment que le SAnMBR a commencé à être utilisé pour le traitement des eaux usées et l’influence de la température et des caractéristiques de la membrane sur l’efficacité du traitement a été étudiée en détail. Cependant, les études sur l’effet tensioactif sur le SAnMBR dans le traitement des eaux usées et sur l’activité des boues pour la production de méthane sont encore limitées.
Récemment, en raison de l’utilisation domestique de détergents ménagers et de lessive, de liquides vaisselle, de shampoings et d’autres produits de soins personnels, une grande quantité de tensioactif anionique, par exemple le sulfonate d’alkylbenzène linéaire (LAS), a été rejetée dans les eaux usées. Cependant, les LAS présents dans les eaux usées ne sont éliminés que par sorption et biodégradation aérobie, mais pas dans des conditions anaérobies en raison de leur caractère hydrophobe et de leurs voies métaboliques restreintes. Si les microbes responsables de la dégradation du LAS existaient dans le réacteur, le LAS adsorbé peut finalement être dégradé après un fonctionnement à long terme. Par conséquent, il est nécessaire de clarifier si le LAS a effectivement été dégradé et converti en méthane ou simplement adsorbé sur les boues lors du traitement des eaux usées par SAnMBR.
En plus de cela, le tensioactif anionique peut également affecter les microbes anaérobies dans le réacteur et l’activité de production de méthane. Il a été rapporté que le LAS favorise un taux de production de méthane plus faible en raison de l’inhibition des étapes méthanogènes et acidogènes du processus anaérobie. La concentration plus élevée de LAS entraîne une production de méthane plus faible, ce qui peut être attribué au taux de croissance plus faible des micro-organismes anaérobies. D’un autre côté, la sorption des tensioactifs entraînera des charges importantes dans les boues d’épuration. Le dodécylsulfate de sodium (SDS) avec une concentration plus faible peut améliorer la fermentation des boues activées en éliminant l’EPS pour augmenter l’efficacité du traitement bactérien ultérieur. Une dose plus élevée inhibera l’activité cellulaire en raison de la structure altérée de la membrane microbienne anaérobie. Étant donné que l’un des avantages du SAnMBR dans le traitement des eaux usées est de récupérer le méthane sous forme de bioénergie, il est d’une grande importance d’explorer l’effet du LAS sur le potentiel méthanogène des boues anaérobies.
Les résultats ont indiqué que le LAS était plutôt adsorbé à la surface des boues que dégradé et que son adsorption entraînait en effet des charges importantes dans les boues d’épuration. L’efficacité de la récupération du méthane dans le traitement des eaux usées a diminué en raison de la présence de LAS dans les eaux usées. Enfin, le mécanisme d’influence du LAS sur le processus anaérobie dans le traitement des eaux usées par SAnMBR a été proposé en fonction de tous les résultats expérimentaux. Les résultats de cette étude devraient fournir des informations utiles sur la question de savoir si le SAnMBR est apte ou non à éliminer les LAS contenant des eaux usées. L’efficacité d’élimination du LAS de 30 à 70 % a été attribuée à l’adsorption plutôt qu’à la dégradation et son adsorption diminue l’activité de production de méthane des boues. Le LAS a eu un effet plus négatif sur les méthanogènes acétoclastiques que le microbiote acidogène et l’inhibition du LAS sur l’activité méthanogène en était responsable.
Présence de tensioactifs dans les eaux usées : variations horaires et saisonnières des eaux usées urbaines et industrielles de Séville
https://www.sciencedirect.com/science/article/abs/pii/S0048969713010486 [20]
La consommation annuelle européenne de tensioactifs en 2011 était de 2,95 Mt, dont 1,40 Mt non ioniques et 1,22 Mt anioniques selon les données rapportées par le Comité européen des tensioactifs organiques et de leurs intermédiaires. Les LAS sont actuellement les tensioactifs anioniques les plus utilisés dans la formulation de détergents ménagers et de lessive, de liquides vaisselle, de shampoings et d’autres produits de soins personnels. Les LAS commerciaux sont disponibles sous forme de mélange d’homologues, avec une longueur différente de chaîne alkyle (de C 10 à 14 ). Les polyéthoxylates de nonylphénol sont des tensioactifs non ioniques utilisés dans les détergents, les peintures, les herbicides, les pesticides, les agents mouillants, les cosmétiques et de nombreux autres produits industriels, agricoles et ménagers.
En raison de leur utilisation et de leur production intensives, ils finissent dans les stations d’épuration des eaux usées, où ils sont éliminés principalement par des processus biologiques et physiques. En raison de voies métaboliques restreintes, les LAS ne sont pas biodégradables dans des conditions anaérobies ; néanmoins, ils sont fortement biodégradés dans des conditions aérobies, mais l’élimination majeure des eaux usées se fait par rétention sur les matières en suspension. Les polyéthoxylates de nonylphénol sont dégradés en intermédiaires métaboliques à chaîne plus courte, notamment le nonylphénol (NP), le mono-nonylphénol et le diéthoxylate.
Les taux d’élimination des LAS étaient supérieurs à 80 % dans la plupart des études rapportées dans la littérature, alors qu’une dispersion plus élevée des taux d’élimination pour NP, NP 1 EO et NP 2 EO (NPE) a été décrit, allant de 0 % à 86 %. Plusieurs auteurs expliquent cette large dispersion par la dégradation des polyéthoxylates de nonylphénol dont les principaux produits de dégradation sont les NPE à chaîne courte. Néanmoins, ces composés ne sont pas complètement éliminés dans les STEP et sont donc continuellement introduits dans l’environnement. Les préoccupations concernant la présence de ces composés dans l’environnement ont conduit à limiter leurs concentrations dans les eaux de surface de l’Union européenne, au moyen de la Directive-cadre sur l’eau 2000/60/CE. Cette directive européenne identifie le NP comme substance prioritaire et fixe sa limite de concentration moyenne annuelle à 0,3 μg L − 1 et sa concentration maximale admissible à 2,0 μg L − 1 dans les eaux de surface.
Évaluation de la communauté microbienne des réacteurs à lit de boues anaérobies à flux ascendant utilisés pour l’élimination et la dégradation du sulfonate d’alkylbenzène linéaire par pyroséquençage
https://www.sciencedirect.com/science/article/abs/pii/S096483051400287X [21]
Les communautés microbiennes de deux réacteurs à lit de boues anaérobies à flux ascendant (UASB) traitant des eaux usées synthétiques complétées par du sulfonate d’alkylbenzène linéaire (LAS) (RS) et des eaux usées de blanchisserie (RL) ont été analysées par pyroséquençage de gènes d’ARNr 16S . Un taux de dégradation du LAS plus élevé a été observé en RL (82 ± 9 %) qu’en RS (45 ± 16 %). Une proportion élevée du taux d’élimination du LAS (55 à 90 %) a été observée dans la région PS pour les deux réacteurs, probablement en raison de la faible concentration de cosubstrats et de la diffusion de l’oxygène dans cette région. Une analyse microbiologique d’échantillons prélevés à 112 jours d’exploitation dans la zone du lit de boues (SB) et de la région du séparateur de phases (PS) des deux réacteurs a confirmé ces résultats. Les communautés microbiennes distinctes trouvées dans chaque réacteur résultant des différentes eaux usées utilisées étaient liées aux taux de dégradation du LAS obtenus. La communauté microbienne du réacteur RL était plus capable de dégrader le LAS, probablement en raison de la présence de xénobiotiques .
L’alkylbenzène sulfonate linéaire (LAS) est largement utilisé à des fins domestiques et industrielles, ce qui conduit à son apparition dans les stations d’épuration des eaux usées à des concentrations allant de 1 à 18 mg.L −1 (Mungray et Kumar, 2009). Des quantités de 1 040 000 t/an de LAS sont produites aux États-Unis, au Japon et en Europe occidentale (Scott et Jones, 2000). La présence de LAS dans les eaux usées pourrait poser problème pour l’application de la technologie anaérobie, car la récalcitrance et l’inhibition du LAS ont été signalées dans les systèmes anaérobies (Garcia et al., 2005). De plus, la présence de LAS dans les milieux aquatiques peut entraîner la formation de mousse dans les ruisseaux et les rivières, dispersant ainsi la pollution. En conséquence, de nombreuses études ont été réalisées pour évaluer la dégradation du LAS dans des conditions anaérobie.
La voie de dégradation du LAS implique des réactions de clivage des cycles aromatiques, de désulfonation et de β- et ω-oxydation dans des conditions aérobies et anaérobies (Lara-Martin et al., 2007). Des organismes aérobies, anaérobies facultatifs et anaérobies seraient impliqués dans la dégradation du LAS. Certains des genres fréquemment signalés comme hébergeant des espèces possédant cette capacité sont Acinetobacter, Aeromonas, Comamonas, Dechloromonas, Desulfovibrio, Geobacter, Holophaga, Parvibaculum, Pseudomonas, Sporomusa, Stenotrophomonas, Variovorax et Zoogloea.
Extraction simultanée et devenir des alkylbenzènesulfonates linéaires, des diéthanolamides de noix de coco, des éthoxylates de nonylphénol et de leurs produits de dégradation dans les stations d’épuration des eaux usées, recevant les eaux côtières et les sédiments de la zone catalane
https://www.sciencedirect.com/science/article/abs/pii/S0021967304013974 [22]
En raison de la formation de métabolites récalcitrants indésirables qui ont donné lieu à des préoccupations environnementales, les APEO sont interdits ou restreints en Europe. La restriction de l’utilisation des APEO dans les détergents ménagers et leur remplacement volontaire dans les formulations industrielles ont entraîné une baisse substantielle des concentrations d’APEO dans toute l’Europe du Nord. Cependant, en Espagne, principalement en raison de coûts de production inférieurs, les APEO sont encore utilisés en quantités importantes dans des applications institutionnelles et industrielles et il est important de continuer à surveiller leur présence et leur distribution dans l’environnement afin de déterminer les tendances de concentration, et donc , prédisent leur sort à long terme.
D’autres tensioactifs ont reçu moins d’attention. Plusieurs études ont porté sur le devenir des alkylbenzène sulfonates linéaires (LAS), des éthoxylates d’alcool (AE) et des diéthanolamides de noix de coco (CDEA) et ont rapporté des niveaux dans les effluents des STEP et dans les eaux côtières. en Catalogne, ce type de contamination reste toutefois à évaluer.
L’élimination des tensioactifs dans certaines STEP catalanes s’est avérée incomplète, ce qui a entraîné des concentrations d’effluents dépassant dans de nombreux cas 20 μg/L pour les composés nonylphénoliques œstrogéniques et 200 μg/L pour les LAS. Cependant, une nette tendance à la baisse des concentrations de composés nonylphénoliques a été observée, en comparant les niveaux actuels et ceux trouvés dans des études antérieures. Cependant, malgré cela, des composés nonylphénoliques sont toujours présents dans le milieu marin recevant les STEP.
Devenir, comportement et effets des tensioactifs et de leurs produits de dégradation dans l’environnement
https://www.sciencedirect.com/science/article/abs/pii/S0160412005001625?via%3Dihub [23]
Les tensioactifs sont largement utilisés dans les produits ménagers et industriels. Après utilisation, les tensioactifs ainsi que leurs produits sont principalement rejetés dans les stations d’épuration puis dispersés dans l’environnement via le rejet des effluents dans les eaux de surface et l’évacuation des boues sur les terrains. Les tensioactifs ont des comportements et des destins différents dans l’environnement. Les tensioactifs non ioniques et cationiques présentaient une sorption sur le sol et les sédiments beaucoup plus élevée que les tensioactifs anioniques tels que le LAS . La plupart des tensioactifs peuvent être dégradés parles microbes présents dans l’environnement, bien que certains tensioactifs tels que le LAS et le DTDMAC ainsi que les alkylphénols puissent être persistants dans des conditions anaérobies. Il a été constaté que les LAS se dégradent dans les sols amendés par des boues avec une demi-vie de 7 à 33 jours. La plupart des surfactants ne sont pas toxiques de manière aiguë pour les organismes aux concentrations environnementales et la toxicité aquatique chronique des surfactants s’est produite à des concentrations généralement supérieures à 0,1 mg/L. Cependant, les alkylphénols se sont révélés capables d’induire la production de vitellogénine chez les poissons mâles à une concentration aussi faible que 5 μg/L. Davantage de données sur la toxicité sont nécessaires pour évaluer les effets sur les organismes terrestres tels que les plantes.
Les tensioactifs constituent un groupe diversifié de produits chimiques conçus pour avoir des propriétés de nettoyage ou de solubilisation. Ils sont généralement constitués d’un groupe de tête polaire (chargé ou non), qui est bien solvaté dans l’eau, et d’une queue d’hydrocarbures non polaires, qui ne se dissout pas facilement dans l’eau. Ainsi, les tensioactifs combinent des propriétés hydrophobes et hydrophiles dans une seule molécule. Les tensioactifs synthétiques sont des produits chimiques économiquement importants. Ils sont largement utilisés dans les détergents ménagers, les produits de soins personnels, les textiles, les peintures, les polymères, les formulations de pesticides, les produits pharmaceutiques, les mines, la récupération du pétrole et les industries des pâtes et papiers. La production mondiale de tensioactifs synthétiques s’élève à 7,2 millions de tonnes par an (Di Corcia, 1998).
Les tensioactifs étaient principalement de trois types : anioniques, non ioniques et cationiques (tableau 1). Les alkylbenzènesulfonates linéaires (LAS), les alkyléthoxysulfates (AES), les alkylsulfates (AS), les alkylphénol éthoxylates (APE), les alkyléthoxylates (AE) et les composés d’ammonium quaternaire (QAC) sont les tensioactifs commerciaux couramment utilisés. En particulier, LAS, APE et QAC sont les tensioactifs les plus étudiés. Dans ce qui suit, nous utilisons des abréviations pour chaque classe de tensioactifs, par exemple C12EO9 (EO = unité oxyde d’éthylène) ayant neuf unités EO et une chaîne alkyle de 12 atomes de carbone, C14LAS ayant une chaîne alkyle de 14 atomes de carbone, NPE9 ou NPEO9. pour les éthoxylates de nonylphénol avec 9 unités EO.
Les alkylbenzènesulfonates linéaires (LAS) sont les tensioactifs anioniques synthétiques les plus couramment utilisés. Il est largement utilisé depuis plus de 30 ans, avec une consommation mondiale estimée à 2,8 millions de tonnes en 1998 (Verge et al., 2000). Les produits disponibles dans le commerce sont des mélanges très complexes contenant des homologues avec des chaînes alkyles allant de 10 à 14 unités carbone (C10-C14LAS). De plus, étant donné que le groupe phényle peut être attaché à n’importe quel atome de carbone interne de la chaîne alkyle, chaque homologue contient 5 à 7 isomères de position.
Les éthoxylates d’alkylphénol (APE) constituent une grande partie du marché des tensioactifs non ioniques. La production mondiale d’APE était estimée à 500 000 tonnes en 1997 avec 80 % d’éthoxylates de nonylphénol (NPE) et 20 % d’éthoxyalyses d’octylphénol (OPE) (Renner, 1997). L’inquiétude s’est récemment accrue quant à l’utilisation généralisée des APE en raison de leurs produits de biodégradation relativement stables, le nonylphénol (NP) et l’octylphénol (OP). Il a été démontré que le NP et l’OP sont toxiques pour les espèces marines et d’eau douce et qu’ils induisent des réponses œstrogéniques chez les poissons.
Les tensioactifs à base d’ammonium quaternaire (QAC) sont des molécules comportant au moins une chaîne hydrocarbonée hydrophobe liée à un atome d’azote chargé positivement, les autres groupes alkyle étant pour la plupart des substituants à chaîne courte tels que les groupes méthyle ou benzyle. Les principales utilisations de ce groupe de tensioactifs cationiques sont les assouplissants textiles et les agents antiseptiques dans les détergents à lessive ainsi que d’autres utilisations industrielles. L’ingrédient actif le plus largement utilisé dans les assouplissants textiles était jusqu’à récemment le chlorure de suif diméthylammonium (DTDMAC). Cependant, le remplacement du DTDMAC par des tensioactifs cationiques esters tels que le chlorure de diéthylester diméthylammonium (DEEDMAC) a récemment commencé en Europe (Giolando et al., 1995).
Après utilisation, les tensioactifs résiduels et leurs produits de dégradation sont rejetés vers les stations d’épuration ou directement dans les eaux de surface, puis dispersés dans différents compartiments environnementaux. En raison de leur utilisation répandue et de leur forte consommation, les tensioactifs et leurs produits de dégradation ont été détectés à diverses concentrations dans les eaux de surface, les sédiments et les sols amendés par des boues. Afin d’évaluer leurs risques environnementaux, nous devons comprendre la distribution, le comportement, le devenir et les effets biologiques de ces tensioactifs dans l’environnement. Il existe des documents de recherche détaillés et des articles de synthèse sur la présence de divers tensioactifs et de leurs produits de dégradation dans l’environnement. Différents types de tensioactifs ont été détectés dans les effluents d’eaux usées avec des concentrations allant jusqu’à 1 090 μg/L pour les LAS, jusqu’à 332 μg/L pour les APE et jusqu’à 62 μg/L pour les APE. μg/L pour le DTDMAC. Ces tensioactifs ont également été trouvés dans les boues traitées à des concentrations élevées allant jusqu’à 30 200 mg/kg de poids sec pour les LAS, jusqu’à 81 mg/kg pour les APE, et jusqu’à à 5 870 mg/kg pour le DTDMAC. Ils ont également été rapportés dans les eaux de surface à des concentrations allant jusqu’à 416 μg/L pour le LAS, et les produits de dégradation de l’APE ont été largement détectés dans différents compartiments environnementaux (air, eau, sédiments). Cet article de synthèse s’est principalement concentré sur le comportement et le devenir ainsi que sur l’écotoxicité de différents types de tensioactifs dans l’environnement.
Les tensioactifs pénétrant dans l’environnement par le rejet d’effluents d’eaux usées dans les eaux de surface et l’épandage de boues d’épuration sur le sol peuvent avoir un impact sur l’écosystème en raison de leur toxicité sur les organismes de l’environnement. Des tensioactifs ont été trouvés dans le milieu aquatique à des niveaux de μg/L, principalement en raison de leur utilisation généralisée et du rejet d’effluents d’eaux usées dans les eaux de surface. Ils sont également mesurés dans les sols amendés par des boues en raison des concentrations résiduelles élevées dans les boues. Les tensioactifs ont une sorption relativement élevée sur les boues, les sédiments et le sol, et leur sorption est de l’ordre : cationique > non ionique > anionique. Ils sont fortement associés aux particules ou aux sédiments.
es tensioactifs pénétrant dans l’environnement par le rejet d’effluents d’eaux usées dans les eaux de surface et l’épandage de boues d’épuration sur le sol peuvent avoir un impact sur l’écosystème en raison de leur toxicité sur les organismes de l’environnement.
Métabolites perturbateurs endocriniens des éthoxylates d’alkylphénol – Un examen critique des méthodes d’analyse, des événements environnementaux, de la toxicité et de la réglementation
https://www.sciencedirect.com/science/article/pii/S0048969718312439?via%3Dihub [24]
Bien que les métabolites des éthoxylates d’alkylphénol (APEO) soient classés comme substances dangereuses, ils continuent d’être rejetés dans l’environnement à partir de diverses sources et ne sont généralement pas surveillés. Leur large utilisation a conduit à une augmentation des voies d’exposition possibles pour l’homme, ce qui est alarmant. De plus, il existe un manque de connaissances sur le comportement de ces métabolites vis-à-vis de l’environnement et de leur toxicité, ainsi que sur leurs effets biologiques sur la santé humaine. L’objectif de ce travail est de donner un aperçu des métabolites de l’APEO et de leur analyse, de leur occurrence et de leur toxicité dans divers échantillons environnementaux et humains. Les métabolites de l’APEO ont des effets néfastes sur les humains, la faune et l’environnement en raison de leur libération dans l’environnement. Actuellement, certaines études sont disponibles sur le comportement des alkylphénols dans le sol, les sédiments, les eaux souterraines, les eaux de surface et les aliments. Cependant, aucun de ces articles ne considère leur toxicité chez l’homme et surtout leur effet sur le système nerveux et immunitaire. Ce travail résume les occurrences environnementales des métabolites des APEO dans des matrices, par exemple l’eau, les aliments et les matrices biologiques, leurs effets sur les systèmes immunitaire et nerveux, ainsi que les problèmes spécifiques aux isomères. Grâce à cet accent, nous sommes en mesure de couvrir les événements les plus courants d’exposition humaine, qu’ils soient directs ou indirects.
1 . Introduction
L’un des principaux groupes de tensioactifs non ioniques sont les éthoxylates de 4-alkylphénol ( APEO ), par exemple les éthoxylates de nonylphénol et d’octylphénol. La capacité de se concentrer sur différentes surfaces et de former des micelles dans des solutions est la principale caractéristique des composés tensioactifs. La structure amphiphile de la molécule tensioactive, qui comporte une partie polaire (hydrophile) et non polaire (hydrophobe), est à l’origine de leur activité de surface. Le caractère hydrophile des APEO est dû à une chaîne polyéthoxylate avec des unités éthoxylates de 1 à 40. La propriété hydrophobe opposée est apportée par les alkylphénols à chaînes hydrocarbonées ramifiées contenant huit, neuf ou douze atomes de carbone. Les APEO les plus courants sont les éthoxylates d’octylphénol ( OPEO ), les éthoxylates de nonylphénol ( NPEO ) et les éthoxylates de dodécylphénol (DOEO).
Les APEO étant des tensioactifs, ils sont largement utilisés dans les formulations de nettoyage et comme auxiliaires de processus industriels. Ils ont donc une large gamme d’applications, notamment comme agents dispersants dans la production de papier et de pâte à papier, agents émulsifiants dans les peintures au latex et les formulations de pesticides, agents de flottation, nettoyants industriels (surfaces métalliques, transformation textile et industrie alimentaire), nettoyants à froid pour voitures, et les produits d’entretien ménager. La plupart des APEO sont utilisés dans des solutions aqueuses pour faciliter leur manipulation, ce qui signifie qu’ils sont rejetés dans les eaux usées municipales et industrielles , puis transportés vers les usines de traitement des eaux usées . Au cours des différentes étapes du traitement des eaux usées, les APEO subissent un processus de biodégradation complexe dans lequel plusieurs micro-organismes métabolisent les APEO par leur chaîne éthoxy et forment plusieurs produits de dégradation , dont les 4-alkylphénols (AP), l’acide (4-alkylphénoxy) acétique ( AP1EC ), Acide (4-alkylphénoxy)éthoxyacétique ( AP2EC ), monoéthoxylate de 4-alkylphénol ( AP1EO ) et diéthoxylate de 4-alkylphénol ( AP2EO ). Ces produits de dégradation sont persistants, bioaccumulables et toxiques. Certaines sont déjà considérées comme des substances prioritaires ou répertoriées comme substances dangereuses prioritaires dans la directive-cadre sur l’eau de l’Union européenne. Parmi ces substances, le NP est considéré comme le métabolite le plus important et le plus dangereux ; les NP et OP les plus courants sont illustrés à la Fig .
Divers métabolites de l’APEO sont omniprésents dans de nombreuses matrices pertinentes pour l’environnement (par exemple, les eaux d’entrée et de sortie des usines de traitement des eaux usées, l’eau des rivières, l’eau de mer, les eaux de surface, les eaux souterraines, l’eau potable , les sédiments et le sol), y compris la nourriture, eau potable et des échantillons de tissus humains, et sont encore plus toxiques que leurs composés d’origine. Outre les propriétés négatives mentionnées, certains métabolites tels que les NP peuvent agir comme l’hormone féminine 17 β -estradiol en se liant au récepteur des œstrogènes et en déplaçant le 17 β -estradiol de manière compétitive.
Étant donné que l’effet œstrogénique et le comportement de dégradation dans l’environnement des APEO individuels et de leurs métabolites dépendent fortement de la structure et de l’encombrement de la chaîne latérale, il est absolument nécessaire de considérer ce problème d’un point de vue spécifique aux isomères. Des études ont montré que le potentiel œstrogénique d’un seul isomère est deux à quatre fois supérieur à celui de la substance de référence, et lorsque l’on compare les isomères biodégradables les plus faciles avec les plus persistants, les demi-vies sont trois à quatre fois plus longues. Les isomères uniques des métabolites de l’APEO sont importants pour évaluer leurs effets écologiques et sanitaires. Cela est vrai même pour leurs énantiomères puisque les systèmes biologiques sont énantiosélectifs.
Une conséquence alarmante de l’utilisation et de la diffusion généralisées des APEO dans l’environnement à travers diverses sources est l’augmentation des voies d’exposition à leurs métabolites pour les humains et la faune sauvage. Ces voies comprennent l’ingestion d’aliments et de produits carnés provenant de terres contaminées ou irriguées avec de l’eau contaminée , et la consommation d’ eau du robinet provenant d’eaux souterraines ou de surfaces polluées. Cependant, l’évaluation des risques ainsi que d’importantes études toxicologiques sur ces substances ont été réalisées sans tenir compte des effets spécifiques aux isomères.
Pour la plupart des métabolites, les effets sur l’homme (notamment sur les systèmes immunitaire et nerveux) et sur l’environnement n’ont pas été suffisamment étudiés. Néanmoins, leur effet sur ces systèmes (par exemple l’homme et l’environnement) pourrait être significatif, c’est pourquoi les métabolites de l’APEO jouent toujours un rôle important dans le débat.
Les grandes lignes de cette revue sont les suivantes : Les premières sections sont consacrées aux méthodes de synthèse et d’analyse des APEO et à la détermination de leurs métabolites. Ceci est suivi d’une discussion de leur présence et de leurs concentrations dans les matrices environnementales et biologiques, y compris les tissus humains. Ensuite, un aperçu de leur toxicité est donné, y compris leurs effets sur la fonction cognitive et leur régulation. Enfin, nous soulignons la nécessité de recherches plus approfondies et résumons les tâches suggérées par les données recueillies ici.
2 . Production, applications et marchés
Le principal composé pour la synthèse des APEO est le phénol. En fonction de l’APEO recherché, il existe trois voies possibles pour la synthèse de ces tensioactifs non ioniques. Dans une alkylation du phénol avec du triméthylpentène, le produit est du 4- tert -octylphénol (OP), dans une réaction catalysée par un acide avec un mélange d’isomères de nonène, le produit est un mélange de 4-nonylphénol (NP), et avec une charge d’oléfine complexe à 70 % Teneur en alkyle en C 12 , le produit est un mélange de 4-dodécylphénol. Ce sont des préproduits pour la synthèse APEO. La synthèse de NP et DP donne déjà un mélange complexe constitué de composés isomères avec des structures diversement ramifiées de leurs chaînes latérales. Les éthoxylates d’alkylphénol sont ensuite synthétisés via une réaction avec de l’oxyde d’éthylène et de l’hydroxyde de potassium/éthanol comme catalyseur. Finalement, cela aboutit à un mélange d’ homologues d’oligomères avec des longueurs variables de chaîne polyéthoxy.
La présence d’APEO et de leurs métabolites dans l’environnement est due à des activités anthropiques. Des analyses approfondies d’échantillons provenant de différents compartiments environnementaux ont été réalisées en ce qui concerne les APEO et leurs métabolites, car ils présentent une très faible biodégradabilité dans les sédiments et sont capables de persister pendant des décennies.
5 . Toxicité
Compte tenu du rôle clé que jouent les œstrogènes dans les processus de développement et les fonctions de reproduction, les composés ayant un effet œstrogénique (c’est-à-dire les métabolites de l’APEO) sont une source de préoccupation majeure. Pour le NP et l’OP, il a été démontré qu’ils sont extrêmement toxiques pour les poissons, les invertébrés et les algues à des concentrations de 17 à 3 000 μg/L et qu’ils présentent une toxicité chronique dans la plage de 3,7 à 6 μg/L. Outre le potentiel œstrogénique déjà bien connu des métabolites de l’APEO, un autre problème important est leur impact sur le développement et le développement du système nerveux et immunitaire des mammifères. Dans la section suivante, nous commençons par une brève discussion du potentiel œstrogénique des métabolites de l’APEO, suivie d’un aperçu de leurs effets sur les systèmes nerveux et immunitaire et d’un bref examen des effets à faible dose. effets des perturbateurs endocriniens sur le système nerveux et immunitaire. Le système immunitaire est un système vulnérable qui peut réagir aux changements par une réaction allergique. Au cours des dernières décennies, les maladies allergiques ont augmenté . Ces maladies complexes dépendent fortement des interactions gènes-environnement et des études épidémiologiques ont montré que divers facteurs de risque peuvent déclencher des maladies allergiques. L’un de ces facteurs de risque est le groupe des perturbateurs endocriniens (EDC), qui peuvent déclencher ou même exacerber des maladies allergiques.
La plupart des études utilisent des concentrations élevées d’alkylphénols pour leurs expériences, ce qui est peu probable en dehors du laboratoire, mais une étude récente a montré que, pour la cellule THP-1 de type macrophage humain, l’OP est capable de déclencher une réponse immunitaire même à de faibles concentrations. (0,001 µM). Dans la société moderne, les produits chimiques font partie de la vie et leur utilisation et leur rejet dans l’environnement ne cessent d’augmenter. Cela inclut les EDC et en particulier les métabolites APEO. Même à faibles doses, les perturbateurs endocriniens peuvent interférer avec le système endocrinien et reproducteur et inhiber la synthèse et/ou le transport d’hormones spécifiques. De plus, les hormones sont un facteur clé dans le bon développement de divers systèmes organiques et tissus tels que ceux de l’appareil reproducteur, du cerveau et du système neuroendocrinien. Les complications de la grossesse telles que l’échec de l’implantation et la perte du fœtus sont quelques-uns des résultats d’un réseau de cytokines déséquilibré à l’interface materno-fœtale. Les perturbateurs endocriniens comme les NP suscitent des inquiétudes considérables quant à l’exposition maternelle pendant la grossesse, même à faibles doses. En 2013, l’Agence européenne des produits chimiques a inclus le NP sur la liste des substances candidates extrêmement préoccupantes à inclure dans l’annexe XIV (Liste d’autorisation) du règlement 1907/2006/CE (REACH).
Fuite de tensioactifs dans les eaux grises : impact environnemental, atténuation et leur économie circulaire
https://www.sciencedirect.com/science/article/abs/pii/S2213343723014549
Rétention de l’arsenic et comportement de transport en présence de tensioactifs anioniques et non ioniques typiques
https://www.sciencedirect.com/science/article/abs/pii/S1001074215004763 [36]
La production massive et l’utilisation généralisée de tensioactifs ont entraîné le rejet d’une grande quantité de résidus de tensioactifs dans l’environnement, ce qui pourrait avoir un impact sur le comportement de l’arsenic. Dans la présente étude, l’influence du tensioactif anionique dodécylbenzène sulfonate de sodium ( SDBS) et du tensioactif non ionique polyéthylèneglycol octylphényléther (Triton X-100) sur le comportement de l’arsenic a été étudiée dans des tests par lots et en colonne. La présence de SDBS et de Triton X-100 a réduit la rétention d’arsenic sur la ferrihydrite (FH), a amélioré le transport de l’arsenic à travers les colonnes de sable enrobé de FH (sable FH) et a favorisé la libération d’arsenic de la surface du FH. Avec des tensioactifs coexistants en solution, la quantité d’arsenic adsorbée à l’équilibre sur le FH a diminué jusqu’à 29,7 % et le taux d’adsorption a diminué jusqu’à 52,3 %. Le pré-revêtement avec des tensioactifs a provoqué une diminution de la quantité adsorbée et du taux d’adsorption de l’arsenic jusqu’à 15,1 % et 58,3 %, respectivement. En raison de l’atténuation de l’adsorption provoquée par les tensioactifs, la percée de l’As (V) et de l’As (III) avec le SDBS dans les colonnes remplies de sable FH était respectivement 23,8 % et 14,3 % plus rapide que celle dans celles sans SDBS. Dans les colonnes remplies de sable FH recouvert de SDBS, le transport de l’arsenic a été davantage amélioré. La percée de l’As(V) et de l’As(III) était 52,4 % et 43,8 % plus rapide et le taux de rétention cumulé était respectivement de 44,5 % et 57,3 % inférieur à celui des systèmes de colonnes de sable FH pur. La mobilisation de l’arsenic par les tensioactifs augmente avec l’augmentation de la quantité initiale d’arsenic adsorbée. La quantité cumulée de libération d’As(V) et d’As(III) de la colonne remplie a atteint respectivement 10,8 % et 36,0 %.
Les tensioactifs sont généralement un groupe de produits chimiques composé de groupes polaires et non polaires. En raison de leurs propriétés de solubilisation et de nettoyage, les tensioactifs sont largement utilisés dans les détergents ménagers, l’industrie textile ainsi que dans les activités de fonderie et d’exploitation minière. La production mondiale de tensioactifs s’élève à 18,2 millions de tonnes par an. Selon les propriétés de leurs groupes polaires, les tensioactifs sont principalement divisés en type anionique, type cationique et type non ionique, parmi lesquels les tensioactifs anioniques et non ioniques sont les plus largement utilisés. Après avoir été utilisés, les résidus de tensioactifs sont rejetés dans les stations d’épuration ou directement dans les milieux environnementaux tels que l’eau, le sol et les sédiments. Cela conduit à la détection de différents niveaux de tensioactifs dans les compartiments environnementaux. La distribution et le comportement des tensioactifs dans l’environnement ont été largement étudiés pour évaluer les risques environnementaux. Lorsqu’ils pénètrent dans l’environnement, les tensioactifs ont tendance à être adsorbés sur les particules et les sédiments, modifiant ainsi les caractéristiques physico-chimiques de la surface des particules et influençant le comportement environnemental d’autres polluants. Il est suggéré que les tensioactifs pourraient exercer des effets significatifs sur l’adsorption et la libération de matières organiques et de métaux lourds. Les mécanismes comprennent principalement la compétition d’adsorption, les réactions de complexation, la solubilisation et la répulsion électrostatique . Compte tenu de l’utilisation répandue de tensioactifs et de la toxicité manifeste de l’arsenic, cette étude a examiné les effets potentiels des tensioactifs sur le comportement de l’arsenic dans l’environnement.
La rétention de l’arsenic sur les particules et son transport dans les sédiments sont deux facteurs principaux contrôlant sa biodisponibilité et sa toxicité. Les deux processus sont dans une large mesure influencés par des substances coexistantes telles que les matières organiques naturelles (MON) et les anions. Il a été bien documenté que ces substances peuvent réduire l’adsorption de l’arsenic et améliorer le transport de l’arsenic par compétition d’adsorption, réactions de complexation, échange d’anions et répulsion électrostatique. Ainsi, grâce à de tels mécanismes, les molécules de tensioactifs peuvent potentiellement influencer le comportement d’adsorption et de transport de l’arsenic dans l’environnement.
Dans les environnements naturels, il pourrait y avoir trois scénarios dans lesquels les tensioactifs influencent l’adsorption et la mobilisation de l’arsenic. Le premier scénario représente des environnements aquatiques où les tensioactifs coexistent avec l’arsenic. Il pourrait y avoir une compétition d’adsorption, car les deux substances sont susceptibles d’être adsorbées par les particules. Le deuxième scénario indique que les eaux usées contenant de l’arsenic pourraient être rejetées dans des eaux et des sédiments recouverts de tensioactifs. Comme on le sait dans des circonstances telles que les sites miniers et les lieux de rejet des eaux usées, de grandes quantités de tensioactifs pourraient être adsorbées sur les sédiments. L’occupation des sites d’adsorption à la surface des particules par des tensioactifs peut entraîner une réduction de l’adsorption de l’arsenic. Dans le troisième scénario, contrairement au deuxième, les sédiments contaminés par l’arsenic pourraient être exposés à des tensioactifs, conduisant à la mobilisation de l’arsenic.
Traitement amélioré par des tensioactifs des sols de la toundra arctique contaminés par le pétrole : évaluation écotoxicologique
https://www.sciencedirect.com/science/article/abs/pii/S2352186421002182?via%3Dihub [37]
Les solutions de tensioactifs et les résidus de tensioactifs dans le sol ont montré un potentiel d’inhibition envers les plantes supérieures. BOK-6, en raison de son profil chimique et écotoxique, a complètement inhibé la germination des graines dans le sol traité. Le tensioactif moins écotoxique AddiMax PV01 a inhibé la croissance des plantes de 38 % par rapport au témoin. Le traitement a augmenté l’écotoxicité et l’activité déshydrogénase activité et a encore abaissé l’ activité catalase du sol de fond et, par conséquent, les résidus de tensioactifs migrant vers les territoires non contaminés doivent être contrôlés. Par cette étude, nous montrons l’efficacité de l’assainissement des sols arctiques grâce aux tensioactifs, mais les risques environnementaux associés doivent être pris en compte.
Le mode d’action du traitement des sols hydrophobes amélioré par des tensioactifs est bien connu ; les avantages de la méthode comprennent l’économie d’eau propre, l’efficacité de la séparation par dispersion, la possibilité d’une utilisation ultérieure des produits pétroliers extraits, la préparation du sol pour une biorestauration ultérieure. Les inconvénients de la méthode sont le risque de pollution secondaire du sol par les substances toxiques et de modification des caractéristiques morphologiques des sols, etc. Le choix des tensioactifs synthétiques pour l’assainissement des sols doit être basé sur différents critères : efficacité, sécurité environnementale et faisabilité économique.
Comparés aux tensioactifs synthétiques, les biosurfactants présentent une activité de surface supérieure malgré leur structure moléculaire volumineuse. D’origine naturelle, les glycolipides (saponines), les lipopeptides, les lipoprotéines, les phospholipides (lécithine), les acides gras, les biosurfactants polymères et particulaires présentent les avantages de biodégradabilité, de production de ressources renouvelables, de régénération possible, de spécificité élevée et de toxicité faible ou non. Leur utilisation dans le traitement des sols a été étudiée par divers chercheurs comme une technologie respectueuse de l’environnement, démontrant une efficacité élevée — jusqu’à 99 %.
Interactions tensioactifs-sol lors de l’assainissement des sols contaminés par des composés organiques hydrophobes modifiés par des tensioactifs
https://www.sciencedirect.com/science/article/abs/pii/S0301479708002399 [38]
Les tensioactifs sont des molécules amphiphiles qui réduisent la tension superficielle aqueuse et augmentent la solubilité des composés organiques hydrophobes (HOC). L’assainissement des sols et aquifères contaminés par des tensioactifs modifiés par des tensioactifs a fait l’objet d’une attention particulière en tant que stratégie de traitement efficace – un concept similaire à l’utilisation de savons et de détergents comme agents de lavage pour éliminer la graisse des tissus souillés. Les mécanismes proposés impliqués dans l’assainissement modifié par les surfactants comprennent : l’abaissement de la tension interfaciale, la solubilisation des HOC par les surfactants et le transfert de phase des HOC de la phase sorbée dans le sol à la phase pseudo-aqueuse. Cependant, comme pour toute contre-mesure chimique proposée , le sort du surfactant ajouté suscite des inquiétudes. Cette revue résume l’état actuel des connaissances concernant la sorption des tensioactifs non ioniques formant des micelles la phase aqueuse , le surfactant sorbé et les phases de surfactant micellaire, ainsi que des modèles récemment développés pour la sorption des surfactants sur le sol. La répartition des HOC est caractérisée par les coefficients apparents de distribution sol-eau en présence de surfactant.sur le sol et sert d’introduction à la recherche sur ce sujet. La sorption du surfactant sur le sol semble augmenter avec l’augmentation de la concentration du surfactant jusqu’au début de la micellisation. Le surfactant en phase sorbée peut représenter la majorité du surfactant ajouté dans les applications d’assainissement modifiés par le surfactant, ce qui peut entraîner une répartition accrue des HOC sur le sol jusqu’à ce que la solubilisation des HOC par le surfactant en phase micellaire entre en concurrence avec succès avec une sorption accrue des HOC sur le sol modifié par le surfactant. Cette revue fournit une discussion sur la théorie du partage à l’équilibre pour tenir compte de la répartition des COH entre le sol.
Les tensioactifs des produits nettoyants polluent dramatiquement l’océan et les littoraux
Performances et réponses de la communauté microbienne de la digestion anaérobie des boues activées résiduaires aux chlorures de benzalkonium résiduels
https://www.sciencedirect.com/science/article/abs/pii/S0196890419312178 [39]
es polluants organiques émergents, les composés d’ammonium quaternaire, ont attiré une grande attention en raison de leur grand enrichissement en boues activées (WAS). Cette étude a sélectionné les chlorures de benzalkonium (BAC) comme modèle pour révéler ses effets sur les niveaux et la longueur des chaînes alkyles (ACL) sur la production de méthane provenant de la digestion anaérobie des WAS . Les résultats ont montré que les BAC provoquaient une libération plus importante de polysaccharides et de protéines solubles, augmentant de 42,6 à 55,7 % et de 11,5 à 19,4 %, ce qui était positivement lié aux niveaux de BAC. En revanche, la méthanogenèse était inhibée et un niveau plus élevé ou un ACL plus court conduisait à une inhibition plus significative. La production maximale de méthane était de 171,1 mL/g VSS en contrôle, et réduite à 97,8, 41,5 et 7,63 mL/g VSS pour ACL16, ACL14 et ACL12 au niveau BAC de 25 mg/g TSS, par conséquent, les accumulations correspondantes de courte durée -les acides gras à chaîne étaient 1,57 à 1,84 fois plus élevés que ceux du contrôle. De plus, la présence de BAC a également affecté la diversité des bactéries et des archées impliquées dans la production de méthane. Par rapport au contrôle, les taux d’alcoolémie ont été réduits les Firmicutes , les Bacteroidetes et les Chloroflexi , en particulier les Firmicutes , avec un rapport de diminution de 12,6 à 20,2 %, tandis que les protéobactéries et les actinobactéries ont augmenté. En outre, un ACL plus long a conduit à un enrichissement plus élevé en méthanomicrobie et, avec la diminution des ACL des BAC de 16 à 12, les méthanogènes dominants sont passés d’acétotrophes à mixotrophes, puis à des méthanogènes hydrogénotrophes. Les résultats de cette étude ont démontré que les niveaux et les ACL des BAC pourraient affecter les performances de la digestion anaérobie des WAS, ainsi que la communauté microbienne.
Le procédé à boues activées s’est avéré être la technologie de base pour traiter les eaux usées. Avec le développement de l’industrialisation et de l’urbanisation, de plus en plus de stations d’épuration des eaux usées ont été exploitées, ce qui entraîne la production de grandes quantités de boues activées WAS. Il a été rapporté que la production annuelle a atteint 4 milliards de tonnes (teneur en humidité de 80 %) en Chine, et qu’environ 80 % d’entre elles n’ont pas été éliminées en toute sécurité. Ces dernières années, le WAS a été considéré comme une source potentielle à la fois d’énergie et de ressources en raison de l’abondance de ses matières organiques et de ses éléments nutritifs et la digestion anaérobie s’est avérée être la méthode préférée et la plus coûteuse. méthode efficace pour récupérer des produits précieux, tels que le méthane, l’hydrogène et les acides gras à chaîne courte (SCFA).
Généralement, la production de méthane à partir de la digestion anaérobie des WAS comprend trois étapes : solubilisation et hydrolyse, acidogenèse et méthanogenèse. À ce jour, en se concentrant sur l’amélioration et l’optimisation de la digestion anaérobie, les facteurs d’influence, tels que le pH, la température, le temps de rétention des boues (SRT) et le rapport carbone/azote, ont été étudiés de manière approfondie. De plus, de nombreuses technologies de prétraitement, notamment les méthodes alcalines, thermiques, ultrasoniques et combinées, ont été explorées. En fait, WAS est également un ensemble d’autres substrats, tels que des métaux lourds, des sulfures et des polluants organiques émergents, qui ont été largement détectés dans les eaux usées et enrichis dans les systèmes de traitement des boues. Parmi eux, le sort des polluants organiques émergents a attiré une grande attention en raison à la fois de leurs variétés et quantités élevées et de leurs grandes menaces potentielles pour les systèmes écosystémiques et la santé des êtres humains. En particulier, les activités des micro-organismes anaérobies peuvent être affectées par les substrats ci-dessus, et le fonctionnement stable du système de digestion anaérobie peut alors être influencé.
Récemment, les composés d’ammonium quaternaire (QAC) ont été fréquemment utilisés comme produits chimiques industriels et domestiques, tels que désinfectants, bactéricides, conservateurs, produits de soins personnels, inhibiteurs de corrosion et auxiliaires technologiques. En conséquence, la plupart des utilisations des CAQ entraînent leur rejet dans l’environnement, et environ 75 % d’entre eux sont enrichis dans les systèmes de traitement des eaux usées, par exemple 0,05 à 6,03 mg/L, 25 à 300 µg/L et 22. –103 mg/kg dans les eaux usées hospitalières, les eaux usées municipales et les boues, respectivement. En particulier, les teneurs en CAQ peuvent atteindre respectivement 4 à 50 mg/L et 4 à 10,5 mg/g de boues sèches dans les systèmes de traitement anaérobie des eaux usées et des boues, et être beaucoup plus élevées dans certains systèmes de traitement des eaux usées industrielles, tels que ceux de l’industrie alimentaire. plantes. Les QAC sont considérés comme des polluants persistants dans l’environnement, ainsi qu’une sorte de contaminants organiques émergents largement enrichis, et sont toxiques pour les organismes aquatiques. De plus, l’exposition à long terme des micro-organismes aux QAC peut entraîner un enrichissement sélectif à la fois des bactéries résistantes aux QAC et des gènes de résistance aux antibiotiques, dont les gènes de multirésistance aux médicaments sont nocifs pour la santé environnementale.
Dans les systèmes aérobies à boues activées, les CAQ peuvent inhiber l’absorption et l’utilisation de l’oxygène [35]. Lorsque la concentration de QACs était inférieure à 120 mg/L, ils peuvent être utilisés comme substrat pour des micro-organismes aérobies, mais dont les activités seraient significativement inhibées au-dessus de 180 mg/L. Bien que les CAQ puissent être biodégradables en unité aérobie, la forte adsorption des CAQ sur les boues activées reste le principal moyen d’élimination lors du traitement des eaux usées. Par exemple, il a été rapporté que 95 % des CAQ étaient adsorbés sur la phase solide dans les systèmes à boues activées. En outre, jusqu’à présent, aucune étude pertinente n’a rapporté que les QAC peuvent être biodégradés dans des conditions anaérobies. Florès et coll et Tezel et al. ont rapporté que lorsque la concentration de QAC dépassait 25 mg/L dans les eaux usées, la méthanogenèse était progressivement inhibée, et elle était complètement inhibée à la concentration de 100 mg/L. Lorsque les QAC fonctionnent comme agents bactéricides pour désintégrer les matières polymères microbiennes, l’efficacité correspondante est substantiellement déterminée par la longueur des chaînes alkyles (ACL) et les niveaux. Cependant, jusqu’à présent, peu d’études ont porté sur les effets des QAC sur la production de méthane provenant de la digestion anaérobie des WAS. De plus, les réponses des micro-organismes anaérobies aux QAC ont rarement été étudiées.
Bioconcentration du tensioactif anionique alkylbenzène sulfonate linéaire (LAS) chez la crevette marine
https://www.sciencedirect.com/science/article/abs/pii/S0025326X14004032 [40]
Dans les régimes d’exposition aiguë, il a été évalué que le LAS agit comme un narcotique polaire et des expositions au LAS dans la plage de 10 à 100 mg L −1 entraîneront une perturbation de la biomembrane. Cette toxicité dépend plutôt de l’efficacité de l’accumulation, de la concentration d’exposition et de la durée de l’exposition au site cible. L’utilisation largement dispersée du LAS, son activité de surface et ses effets bio-membranaires, ainsi que son rejet dans les eaux côtières, souvent sans traitement des eaux usées, sont autant de raisons pertinentes pour étudier sa toxicocinétique et sa distribution dans les organismes marins. Cette évaluation représente une étape clé pour dériver les valeurs du FBC, estimer la CSEO et les valeurs de charge corporelle critique utiles dans les évaluations des risques.
Approche analytique des agents tensioactifs dans l’environnement et enjeux
https://www.sciencedirect.com/science/article/abs/pii/S2214158818300230 [28]
Les tensioactifs sont un composant anthropique important de l’environnement aquatique, utilisés dans un grand nombre de produits ménagers et industriels et sont souvent rejetés dans les réservoirs d’eau douce. L’utilisation croissante des tensioactifs dans la vie quotidienne et dans les industries a conduit à surveiller leurs niveaux et leur impact toxique sur l’environnement. Ces tensioactifs subissent différents types de modifications physiques et chimiques lorsqu’ils sont rejetés dans l’environnement. La présence de tensioactifs dans l’eau peut entraîner la mort de bactéries, champignons et autres micro-organismes. Des concentrations plus élevées peuvent avoir des effets sur la santé humaine, comme une irritation de la peau, des yeux et du système respiratoire [9]. L’effet toxique des tensioactifs dépend de leur activité de surface. L’interaction des tensioactifs avec la membrane cellulaire du biote dépend des activités de surface, car l’interaction avec la membrane est facilitée par la tension superficielle. Le tableau 2 montre la toxicité des tensioactifs (CL 50 et EC 50 ), qui détermine l’étendue de leur applicabilité, comme l’ont rapporté de nombreux chercheurs. Les tensioactifs sont également responsables des processus chimiques dans les eaux des rivières et des effluents liquides des stations d’épuration et de la réduction des paramètres de qualité de l’eau. Les propriétés amphiphiles de ces tensioactifs permettent leur transport en dépôt sec et humide vers les eaux de surface et de ruissellement. Les interactions physiques possibles des molécules amphiphiles dans un environnement aqueux sont illustrées sur la figure 3. De plus, la réduction de la tension superficielle permet aux autres polluants de s’écouler facilement et fournit également des conditions appropriées aux organismes vivants dans l’eau. Les eaux de surface et les eaux usées sont les principales sources de tensioactifs dans l’environnement. Les eaux souterraines peuvent également contenir une petite quantité de tensioactifs en raison du lessivage des eaux usées contaminées. Il est maintenant établi que ces substances chimiques pénètrent dans l’environnement à partir d’un certain nombre de sources et de voies, par le biais des effluents d’eaux usées provenant des usines de traitement municipales, des fosses septiques, des effluents des hôpitaux, des activités d’élevage, des déchets agricoles et industriels. Par conséquent, il existe un besoin urgent d’une compréhension globale de la chimie des voies et des charges environnementales, ainsi que des protocoles AQ/CQ pour l’analyse d’échantillons provenant de différentes sources afin d’utiliser en toute sécurité les tensioactifs.
Dans la présente revue, les principes fondamentaux des tensioactifs et leurs différents aspects dans les méthodes analytiques instrumentales, d’extraction et de validation analytique sont concentrés. De plus, la concentration de tensioactifs rapportée dans des échantillons solides, liquides et aériens sur différents sites dans le monde a été intégrée. Les lacunes actuelles dans notre compréhension concernant l’analyse des tensioactifs dans divers échantillons solides et liquides sont mises en évidence. Il a été établi à plusieurs reprises que les parties hydrophobes et hydrophiles interagissent facilement avec les sous-structures polaires et apolaires des marcomolécules telles que les protéines. Les tensioactifs anioniques eux-mêmes présentent également une activité biologique marquée, soit en se liant à diverses macromolécules bioactives telles que l’amidon, les protéines, les peptides et l’ADN, soit en insérant dans divers fragments cellulaires (c’est-à-dire les membranes phospholipidiques) provoquant un dysfonctionnement. Les tensioactifs anioniques peuvent influencer considérablement l’efficacité biologique des ingrédients actifs dans les formulations pharmaceutiques, soit en se liant directement au médicament, soit en influençant les processus d’adsorption et d’absorption et la répartition des médicaments entre les substances hydrophobes.
Présence et risque environnemental potentiel des tensioactifs et de leurs produits de transformation rejetés par les stations d’épuration des eaux usées
https://www.sciencedirect.com/science/article/abs/pii/S0048969719320030 [27]
En raison de la multitude de produits chimiques présents dans les milieux aquatiques, de nouvelles méthodes de priorisation ont évolué ces dernières années pour concentrer les efforts sur les substances présentant la plus haute priorité d’atténuation. Ces méthodes appliquent souvent des indicateurs liés à l’occurrence, au danger et au risque de composés uniques, l’accent étant mis particulièrement sur ces derniers dans des contextes environnementaux.
Des échantillons d’effluents composites de sept jours provenant d’une campagne de surveillance allemande incluant 33 stations d’épuration conventionnelles (STEP) ont été analysés pour détecter les alkylbenzènesulfonates linéaires ( LAS) et les alkyléthoxysulfates (AES) et ont été examinés par une sélection de suspects à grande échelle pour 1564 tensioactifs et leur transformation. produits(TP) par UHPLC-ESI-QTOF-MS. Les échantillons d’influents composites correspondants sur sept jours de stations d’épuration sélectionnées ont montré des concentrations d’influents élevées ainsi que des taux d’élimination très élevés pour le LAS et l’AES. Cependant, les concentrations moyennes totales des effluents LAS et AES étaient toujours de 14,4 μg/L et 0,57 μg/L, respectivement. Les sous-produits LAS, les di-alkyltétraline sulfonates (DATS), les TP sulfophényl alkyle acides sulfophényl alkyl carboxyliques(SPAC) et les acides sulfo-tétraline alkylcarboxyliques (STAC) ont atteint des concentrations maximales dans les effluents de 19 μg/L, 17 μg/L et 5,3 μg/L, respectivement. Dans de nombreux cas, la somme des concentrations de tous les sous-produits et TP liés aux LAS dépassait la concentration des précurseurs. Des concentrations élevées allant jusqu’à 7,4 μg/L ont été trouvées pour 41 homologues de polyéthylèneglycol. Les tensioactifs quantifiés, leurs TP et leurs sous-produits représentaient ensemble des concentrations allant jusqu’à 82 μg/L dans les effluents des STEP. Pour déterminer le risque de tensioactifs individuels et de leurs mélanges, des homologues uniques ont été regroupés selon une « approche pondérée du nombre de carbones » pour dériver des concentrations estimées sans effet (PNEC) normalisées, sur la base de données expérimentales d’écotoxicité issues d’évaluations de risques existantes, complétées par une structure quantitative appropriée. -Prédictions des relations d’activité (QSAR). Les concentrations environnementales estimées (CEE) ont été calculées en divisant les concentrations de tensioactifs dans les effluents par des facteurs de dilution locaux. Les risques pour tous les tensioactifs analysés étaient inférieurs au rapport PEC/PNEC communément accepté de 1 pour des composés uniques, tandis que les contributions aux effets toxiques du mélange provenant des niveaux de fond de LAS et de DATS ne peuvent être exclues. Les concentrations maximales de LAS dépassaient la moitié de sa PNEC, ce qui pourrait déclencher un examen préalable à l’échelle nationale pour enquêter sur les risques environnementaux potentiels.
Approche analytique des agents tensioactifs dans l’environnement et enjeux
https://www.sciencedirect.com/science/article/abs/pii/S2214158818300230 [25]
Les tensioactifs zwittérioniques, qui présentent les caractéristiques des tensioactifs à la fois anioniques et cationiques, par exemple le méthyldodécyleaminepropanesulfonate (MDAS), sont largement utilisés dans les détergents et l’industrie du moussage et ont un effet toxique néfaste sur l’environnement. Les tensioactifs Gemini, un tensioactif synthétique, deviennent également populaires car ils possèdent des caractéristiques uniques et une efficacité plus élevée pour réduire la tension superficielle de l’interface huile-eau et réduisent ainsi les effets toxiques pour l’environnement.
Les tensioactifs sont un composant anthropique important de l’environnement aquatique, utilisés dans un grand nombre de produits ménagers et industriels et sont souvent rejetés dans les réservoirs d’eau douce. L’utilisation croissante des tensioactifs dans la vie quotidienne et dans les industries a conduit à surveiller leurs niveaux et leur impact toxique sur l’environnement. Ces tensioactifs subissent différents types de modifications physiques et chimiques lorsqu’ils sont rejetés dans l’environnement. La présence de tensioactifs dans l’eau peut entraîner la mort de bactéries, champignons et autres micro-organismes. Des concentrations plus élevées peuvent avoir des effets sur la santé humaine, comme une irritation de la peau, des yeux et du système respiratoire [9]. L’effet toxique des tensioactifs dépend de leur activité de surface. L’interaction des tensioactifs avec la membrane cellulaire du biote dépend des activités de surface, car l’interaction avec la membrane est facilitée par la tension superficielle. Les tensioactifs sont également responsables des processus chimiques dans les eaux des rivières et des effluents liquides des stations d’épuration et de la réduction des paramètres de qualité de l’eau. Les propriétés amphiphiles de ces tensioactifs permettent leur transport en dépôt sec et humide vers les eaux de surface et de ruissellement. De plus, la réduction de la tension superficielle permet aux autres polluants de s’écouler facilement et fournit également des conditions appropriées aux organismes vivants dans l’eau. Les eaux de surface et les eaux usées sont les principales sources de tensioactifs dans l’environnement. Les eaux souterraines peuvent également contenir une petite quantité de tensioactifs en raison du lessivage des eaux usées contaminées. Il est maintenant établi que ces substances chimiques pénètrent dans l’environnement à partir d’un certain nombre de sources et de voies, par le biais des effluents d’eaux usées provenant des usines de traitement municipales, des fosses septiques, des effluents des hôpitaux, des activités d’élevage, des déchets agricoles et industriels. Par conséquent, il existe un besoin urgent d’une compréhension globale de la chimie des voies et des charges environnementales, ainsi que des protocoles AQ/CQ pour l’analyse d’échantillons provenant de différentes sources afin d’utiliser en toute sécurité les tensioactifs.
Activité biologique et impact environnemental des tensioactifs anioniques
https://www.sciencedirect.com/science/article/abs/pii/S0160412002000326 [26]
Les paramètres chimiques et physicochimiques liés à l’activité biologique et au domaine d’application sont brièvement abordés. Des exemples sur l’effet des tensioactifs anioniques sur les membranes cellulaires, sur l’activité des enzymes, sur la liaison à diverses protéines et à d’autres composants cellulaires et sur leur toxicité pour l’homme sont présentés et le mode d’action possible est élucidé. Les sources de pollution environnementale causées par les tensioactifs anioniques sont répertoriées et les méthodes développées pour leur élimination des matrices liquides, semi-liquides et solides sont collectées. Les effets bénéfiques et néfastes des tensioactifs anioniques sur l’environnement sont rapportés et discutés de manière critique. Il a été conclu que le rôle des tensioactifs anioniques dans l’environnement est ambigu : ils peuvent provoquer une grave pollution de l’environnement ayant un effet toxique sur les organismes vivants ; sinon, ils peuvent favoriser la décomposition et/ou l’élimination d’autres polluants inorganiques et organiques de l’environnement. La relation entre leur structure chimique, leurs paramètres physico-chimiques, leur activité biologique et leur impact environnemental n’est pas bien comprise. Un nombre considérable de données sont nécessaires pour le développement de nouveaux tensioactifs anioniques et pour l’application réussie de ceux existants afin de réduire l’adversaire et de promouvoir les effets bénéfiques.
Les effets des lipides et des détergents sur la glycoprotéine P ATPase-active
https://www.sciencedirect.com/science/article/abs/pii/0005273693903392 [29]
Nous avons précédemment isolé et caractérisé une préparation partiellement purifiée de glycoprotéine P active par ATPase, le transporteur multidrogue. L’effet de divers détergents et phospholipides membranaires sur l’activité ATPase de la glycoprotéine P a maintenant été étudié. L’activité de la P-glycoprotéine ATPase était la plus stable dans CHAPS, avec plus de 50 % de l’activité conservée à une concentration de 8 mM. L’octyle glucoside dans la plage des mM faibles a également soutenu l’ATPase, tandis que le désoxycholate a détruit toute activité à 1 mM. La digitonine et le SDS ont inhibé l’activité ATPase à de très faibles concentrations. Le Triton X-100 à 2-10 μM a stimulé l’ATPase presque 2 fois, tandis que des niveaux plus élevés ont inhibé l’activité. Bien que la glycoprotéine P ATPase soit sensible à l’inactivation thermique, sa pleine activité est préservée en présence d’asolectine, mais pas d’espèces de phosphatidylcholine. D’autres études ont révélé que l’asolectine, les phosphatidyléthanolamines saturées et insaturées et la phosphatidylsérine, étaient les mieux à même de maintenir l’activité ATPase à 23°C. Les espèces saturées de phosphatidyléthanolamine ont activé la glycoprotéine P ATPase jusqu’à 40 % à 23 °C et 80 % à 4 °C. Après une délipidation au détergent, divers lipides ont pu restaurer l’activité ATPase de la glycoprotéine P. La phosphatidylcholine insaturée et la phosphatidylsérine étaient les plus efficaces, tandis que les espèces saturées n’étaient pas capables de restaurer l’activité catalytique. Ces résultats indiquent que les lipides membranaires sont nécessaires à l’activité catalytique des domaines ATPase de la glycoprotéine P. La glycoprotéine P a des préférences lipidiques bien définies, les phosphatidyléthanolamines saturées activant à la fois l’ATPase et offrant une protection contre l’inactivation thermique, tandis que les mélanges lipidiques fluides sont capables de restaurer l’activité après la délipidation.
Les protéines de transport sont essentielles aux cellules car elles permettent l’échange de substances entre les cellules et leur environnement à travers la bicouche lipidique formant une barrière étanche. Les lipides membranaires modulent la fonction des protéines transmembranaires telles que les transporteurs de deux manières : les lipides sont étroitement et spécifiquement liés aux protéines de transport et, en outre, ils modulent à partir de la majeure partie de la bicouche lipidique la fonction des protéines de transport. Cet aperçu résume les informations actuellement disponibles au niveau ultra structural sur les lipides étroitement liés aux protéines de transport et l’impact de la modification de la composition lipidique de la membrane globale. Les maladies humaines entraînant une altération de l’homéostasie lipidique entraîneront une altération de la composition lipidique membranaire, qui à son tour affectera la fonction des protéines transporteuses.
Médicamenteux
Utilisation d’antibiotiques en aquaculture et impact sur les eaux côtières
https://www.sciencedirect.com/science/article/abs/pii/S0025326X23000760
Produits pharmaceutiques psychiatriques dans l’environnement
https://www.sciencedirect.com/science/article/abs/pii/S0045653509010832
Les produits pharmaceutiques constituent un groupe vaste et diversifié de composés organiques utilisés en très grandes quantités dans le monde. Aujourd’hui et dans l’Union européenne, il existe plus de 3 000 substances actives sur le marché. Les premiers rapports connus faisant spécifiquement référence à l’élimination incomplète de certains produits pharmaceutiques par le traitement des eaux usées et à leur rejet dans l’environnement par les stations d’épuration des eaux usées (STEP) ont été publiés dans les années 1960 et 1970. Malgré ces premières découvertes indiquant que les produits pharmaceutiques constituent un groupe potentiel de contaminants environnementaux, cette question n’a retenu l’attention que dans les années 1990, lorsqu’elle a été découverte. que certains composés ont la capacité d’interférer avec les écosystèmes à des concentrations aussi faibles que quelques nanogrammes par litre. C’est également au cours de cette décennie que furent développées les premières méthodes analytiques optimisées pour la quantification de produits pharmaceutiques dans des échantillons environnementaux, permettant la détermination de très petites quantités dans les milieux aquatiques ). Depuis, suite à plusieurs études publiées faisant état de la présence de ces composés dans l’environnement, ainsi que les grandes quantités de produits pharmaceutiques produits ainsi que leur utilisation et leur diversité croissantes , l’existence de médicaments pharmaceutiques et de métabolites pharmaceutiquement actifs dans l’environnement a été considérée comme l’une des préoccupations émergentes en matière d’environnement. sciences. Malgré le nombre croissant d’études publiées sur ce sujet ces dernières années, il reste encore beaucoup à comprendre sur les transformations environnementales, leur devenir. En outre, et compte tenu du fait que les produits pharmaceutiques ne sont pas présents dans l’environnement individuellement mais sous forme de mélanges complexes, plusieurs études ont montré que la toxicité des produits pharmaceutiques pour les organismes non ciblés peut se produire à des concentrations pertinentes pour l’environnement en raison d’effets combinés et synergiques
Jusqu’à présent, une grande diversité de produits pharmaceutiques a été trouvée dans l’environnement : analgésiques, antibiotiques, antiépileptiques, β-bloquants, régulateurs lipidiques sanguins, antidépresseurs, anxiolytiques, sédatifs, contraceptifs, etc. Cet article sera spécifiquement axé sur les médicaments psychiatriques (comprenant les anxiolytiques, les sédatifs, les hypnotiques, les antidépresseurs – les inhibiteurs sélectifs de la recapture de la sérotonine, les antidépresseurs tricycliques et autres). Les médicaments psychiatriques constituent un groupe de produits pharmaceutiques couramment prescrits.
Les produits pharmaceutiques sont utilisés dans le but d’avoir un certain type d’effet biologique ou physiologique chez les humains ou les animaux. Parmi leurs caractéristiques spécifiques, ces composés ont la capacité de traverser les membranes cellulaires et sont relativement persistants afin de ne pas être inactivés avant d’avoir l’effet thérapeutique souhaité. Dans le cas particulier des produits pharmaceutiques liés au système nerveux, et en plus des propriétés intrinsèques mentionnées, ceux-ci ont une grande importance pour la régulation du comportement, ayant la capacité d’affecter directement le système nerveux central et de perturber la signalisation neuro-endocrinienne. L’altération des modes de reproduction chez les organismes aquatiques non ciblés est un bon exemple illustrant les effets néfastes possibles lors des tests. organismes, reflétant ainsi le mode d’action de ce groupe particulier de produits pharmaceutiques. Plusieurs études ont démontré que ces composés peuvent affecter les systèmes physiologiques à de très faibles concentrations. van der Ven et coll. (2006) ont démontré que la miansérine, un antidépresseur tétracyclique, possède une activité œstrogénique et produit une perturbation endocrinienne chez le poisson zèbre. Un certain nombre d’autres études sur la fluoxétine, le diazépam, la sertraline, la paroxétine et d’autres ont clairement montré des effets indésirables significatifs des antidépresseurs et des anxiolytiques sur les organismes vivants des matrices aquatiques.
À l’instar d’autres groupes de produits pharmaceutiques, les médicaments psychiatriques ne sont pas complètement métabolisés par le corps humain et sont excrétés sous forme de composé parent inchangé ou sous forme de métabolites ou de conjugués (généralement des glucuronides). Même si les produits pharmaceutiques sont largement métabolisés, leurs métabolites peuvent continuer à être biologiquement actifs ou, dans certains cas, être facilement transformés en composé d’origine dans des conditions environnementales, en raison de l’action bactérienne . De préférence, les produits pharmaceutiques pénètrent dans l’environnement par les effluents des stations d’épuration et l’épandage des boues, en raison de l’insuffisance des méthodes de traitement d’élimination. En outre, ces composés peuvent atteindre les environnements aquatiques et terrestres non seulement en raison de leur utilisation à des fins thérapeutiques humaines, mais également en raison de leur utilisation dans les traitements vétérinaires, des rejets des industries de production, de l’utilisation d’eaux usées traitées pour l’irrigation et une élimination incorrecte par les ménages. En 2005, Bound et ses collaborateurs ont mené une enquête auprès d’environ 400 ménages du sud-est de l’Angleterre sur l’élimination et la perception des risques des produits pharmaceutiques pour l’environnement. Leur enquête a révélé que les antidépresseurs, parmi les huit groupes pharmacologiques ciblés, sont perçus comme l’un des groupes les plus dangereux pour l’environnement par 64,9 % des personnes interrogées. Même si aucune des personnes interrogées n’a admis avoir jeté les antidépresseurs non utilisés dans l’évier, 66,7 % d’entre elles ont jeté les paquets non utilisés à la poubelle avec les résidus organiques domestiques courants. Cependant, l’excrétion par les patients est considérée comme une source de contamination pharmaceutique plus importante que l’élimination incorrecte des emballages inutilisés
Composés pharmaceutiquement actifs et produits chimiques perturbateurs endocriniens dans l’eau, les sédiments et les mollusques des écosystèmes de mangrove de Singapour
https://www.sciencedirect.com/science/article/abs/pii/S0025326X16305100
Cette étude a examiné la présence de bisphénol A (BPA), d’atrazine et de composés pharmaceutiquement actifs sélectionnés (PhAC) dans les habitats de mangrove de Singapour en 2012-2013, à l’aide de plusieurs outils (échantillonnage de sédiments, POCIS et mollusques filtreurs ). Grâce au POCIS, la même série de contaminants (atrazine, BPA et onze PhAC) a été détectée dans les eaux de mangrove au cours de déploiements de 28 jours en 2012 et 2013. Les concentrations de POCIS variaient de pg/L à μg/L. La caféine, le BPA, la carbamazépine, l’E1, le triclosan , la sulfamérazine , la sulfaméthazine etla lincomycineont également été détectés dans les sédiments de mangrove, du faible pg/g de poids sec (par exemple carbamazépine) à ng/g de poids sec (par exemple BPA). La détection de caféine, de carbamazépine, de BPA,de sulfaméthoxazole ou de lincomycinedans les tissus des bivalves a également montré que ces produits chimiques sont biodisponibles dans l’habitat des mangroves. Étant donné que certaines indications suggèrent que certaines substances pharmaceutiquement actives pourraient être biologiquement actives dans la plage de faibles ppb chez les espèces marines, une évaluation plus approfondie devrait être réalisée sur la base de données écotoxicologiques spécifiques aux espèces de mangroves.
Des produits pharmaceutiques humains et divers perturbateurs endocriniens (EDC), résidus d’eaux usées brutes et traitées, ont été détectés dans les eaux de surface, les eaux souterraines et les sols. Des résidus pharmaceutiques ont également été détectés dans les eaux côtières. En fait, même si une fraction importante de la population mondiale vit dans des villes côtières, on a accordé beaucoup moins d’attention à la compréhension de la libération de produits pharmaceutiques dans les eaux côtières et de leurs impacts ultérieurs sur les écosystèmes marins.
Les écosystèmes de mangroves sont des écosystèmes côtiers de transition uniques, généralement confinés aux régions tropicales et subtropicales. Les forêts de mangroves disparaissent rapidement, soit à cause du défrichement direct, soit parce qu’elles sont vulnérables aux facteurs de stress (par exemple élévation du niveau de la mer, infestation d’insectes, pollution chimique). En ce qui concerne la pollution chimique, Bayen (2012) a noté que très peu d’informations sont disponibles sur la présence de composés autres que les composés aromatiques polycycliques, les polluants organiques persistants (POP) conventionnels et les métaux traces. En ce qui concerne les contaminants émergents tels que les résidus pharmaceutiques, la situation n’a pas beaucoup évolué depuis 2012, et seuls quelques-uns ont étudié le comportement et la présence de ces contaminants dans les écosystèmes de mangrove. Ramaswamy et coll. (2011) avaient signalé les niveaux de carbamazépine (un anticonvulsivant et antiépileptique) et de triclosan (un agent antimicrobien) dans les eaux de mangrove en Inde, provenant probablement des eaux usées domestiques et des effluents industriels. Froehner et coll. (2011) ont détecté la présence d’estrone (E1), de 17-β-estradiol et de 17-α-éthynilestradiol dans des sédiments de mangrove au Brésil, associés à des rejets irréguliers ou illégaux d’effluents. Plus récemment, Beretta et al. (2014) ont identifié divers produits pharmaceutiques et de soins personnels dans l’une des plus grandes baies du Brésil, y compris certains sites de mangroves.
Effets potentiels génotoxiques et immunotoxiques de certains médicaments psychotropes et antibiotiques sur les hémocytes de la moule bleue
https://www.sciencedirect.com/science/article/abs/pii/S0269749115001475
La toxicité potentielle des produits pharmaceutiques envers les invertébrés aquatiques est encore mal comprise et parfois controversée. Cette étude vise à documenter la génotoxicité et l’immunotoxicité in vitro des médicaments psychotropes et des antibiotiques sur Mytilus edulis. Les hémocytes de moules ont été exposés à la fluoxétine, à la paroxétine, à la venlafaxine, à la carbamazépine, au sulfaméthoxazole, au triméthoprime et à l’érythromycine, à des concentrations allant de μg/L à mg/L. La paroxétine à 1,5 μg/L a entraîné des dommages à l’ADN tandis que la même concentration de venlafaxine a provoqué une immunomodulation. L’exposition à la fluoxétine a entraîné une génotoxicité, une immunotoxicité et une cytotoxicité. Dans le cas des antibiotiques, le triméthoprime était génotoxique à 200 μg/L et immunotoxique à 20 mg/L alors que l’érythromycine provoquait les mêmes effets néfastes à des concentrations plus élevées. Le métabolisme de l’ADN semble être une cible très sensible pour les médicaments psychotropes et les antibiotiques. De plus, ces composés affectent le système immunitaire des bivalves, avec une intensité variable. Ceci atteste de la pertinence de ces critères pour évaluer le mode d’action toxique des produits pharmaceutiques en milieu aquatique.
Modèles de distribution des polluants organiques et des processus microbiens dans les sédiments marins selon un gradient d’impact anthropique
https://www.sciencedirect.com/science/article/abs/pii/S0269749118311084
Les sédiments marins font partie du cycle hydrologique et constituent le compartiment de stockage ultime de la matière organique d’origine terrestre, y compris des polluants. Étant donné que les processus microbiens pertinents se produisant au niveau benthique peuvent affecter la qualité du système aquatique dans son ensemble, la nécessité d’incorporer des informations sur ledes communautés microbiennespour la modélisation des écosystèmes se fait sentir. Le but de cette étude de terrain était d’explorer les liens existant entre les modèles de contamination des sédiments par trois classes sélectionnées de polluants organiques (HAP, nonylphénols, NP, bisphénol A, BPA) et les principales propriétés microbiennes à travers un gradient de pollution anthropique. Les sédiments ont été échantillonnés sur 34 sites sélectionnés le long de 700 km de la côte ouest de la mer Adriatique. La contamination organique était modérée et a diminué vers le sud. La quantité de carbone associé aux HAP (C-PAH) a augmenté de manière significative avec le carbone organique (OC) des sédiments, ainsi que les taux fonctionnels microbiens. La relation négative entre le rapport PCP/CR et l’OC indique une évolution vers des processus oxydatifs en réponse à la pollution organique et à la toxicité potentielle, estimée en équivalents toxiques (TEQ). Nos résultats ont montré que la contamination organique des sédiments et les processus microbiens benthiques peuvent être intimement liés, avec des répercussions potentielles sur les taux d’émission de CO2et le cycle du C au sein du réseau trophique basé sur les détritus.
Les sédiments marins représentent le compartiment de stockage ultime de tous les matériaux transportés des continents vers les océans, agissant essentiellement comme un puits pour la matière organique, les nutriments et les apports anthropiques, y compris les produits chimiques dangereux.
La directive 2013/39/UE, modifiant les directives 2000/60/CE et 2008/105/CE, rend compte des normes de qualité environnementale (NQE) pour une longue liste de substances chimiques prioritaires et des procédures de mesure de leurs concentrations dans les sédiments marins. Les polluants organiques persistants, très préjudiciables à la santé des organismes, qui peuvent se bio/accumuler dans les sédiments et les organismes en raison de leur hydrophobicité et de leur lipophilie inhérentes sont particulièrement préoccupants. Il est également alarmant de constater que le biote benthique comestible peut être contaminé par des polluants organiques dans les systèmes côtiers marins.
Trois groupes clés de substances organiques persistantes sont les HAPs, les nonylphénols (NP) et le bisphénol A (BPA, 2,2-bis(4-hydroxyphényl)propane). Les HAP comprennent plusieurs centaines de composés organiques chimiquement apparentés et de structures diverses. Ces contaminants, générés principalement lors de la combustion incomplète de matières organiques, sont détectés partout dans le monde, y compris dans les zones reculées (Fernández et al., 2003). Les HAP sont des contaminants potentiellement cancérigènes, mutagènes et toxiques et leur toxicité sur les invertébrés aquatiques est décrite. Les NP comprennent les dérivés métaboliques des tensioactifs non ioniques, les nonylphénoléthoxylates (NPEOn), utilisés comme agents émulsifiants dans les pesticides, additifs pour huiles lubrifiantes, catalyseurs dans le durcissement des résines époxy, dans les blanchisseries industrielles. Le composé le plus produit et mesuré dans l’environnement est le 4-nonylphénol (4-NP), qui est un agoniste œstrogénique hautement toxique pour les poissons, les invertébrés aquatiques et les plantes aquatiques. Le BPA est répertorié comme perturbateur endocrinien et son utilisation est interdite dans la fabrication des biberons (Directive 2011/8/UE) et réduite dans les jouets (Directive 2014/81/UE). Bien que l’activité œstrogénique soit bien connue, la production de BPA a connu une croissance constante ces dernières années, compte tenu de la demande croissante de polycarbonates et de résines époxy. Au cours de la production, des opérations de transport et de l’élimination dans les décharges, le BPA peut être rejeté dans l’environnement à partir de nombreux produits, eaux usées et boues d’épuration.
Les contaminants organiques peuvent atteindre les eaux de surface de différentes manières, notamment par dépôt atmosphérique, ruissellement urbain, effluents municipaux et industriels, déversements ou fuites de pétrole. En raison de leur faible solubilité aqueuse et de leur forte nature hydrophobe, ils ont tendance à être associés aux particules présentes dans le milieu aquatique, les sédiments sous-jacents étant leurs récepteurs ultimes. Spécifiquement pour les sédiments marins contaminés, la Directive 2013/39/UE indique les NQE de différents isomères de HAP, tels que le Benzo(a)pyrène (>30 ng g −1 ), le Benzo(b)fluoranthène (>40 ng g −1 ) , Benzo(k)fluoranthène (>20 ng g −1 ), Benzo(g,h,i)pérylène (>55 ng g −1 ), Indenopyrène (>70 ng g −1 ), Anthracène (>24 ng g −1 ), Fluoranthène (>110 ng g −1 ) et Naphtalène (>35 ng g −1 ). Les NQE pour les NP et le BPA n’ont pas été établies pour les sédiments marins, bien que les Recommandations canadiennes pour la qualité suggèrent des limites de 1,0 mg kg −1 pour les NP, exprimées en unités équivalentes toxiques de nonylphénol, et de 25 μg kg −1 pour le BPA (CCME, 2007; LCPE , 2018). La Commission européenne laisse aux États membres le soin d’identifier sur quelle matrice (eau, sédiment ou biote) les NQE doivent être appliquées. Bien que les poissons, les crustacés , les céphalopodes et les bivalves soient explicitement pris en compte dans la réglementation actuelle, le riche biote des sédiments comprend fondamentalement des communautés microbiennes, responsables du recyclage des nutriments et de la dégradation des composés organiques et des polluants. Au niveau benthique, des processus microbiens pertinents se produisent et peuvent affecter la qualité globale de l’eau. Étonnamment, aucun des descripteurs liés aux microbes n’a été pris en compte dans aucune des directives européennes. La nécessité d’intégrer des informations sur les communautés microbiennes dans l’évaluation du fonctionnement des écosystèmes se fait désormais sentir. Les données de terrain explorant les modèles de contamination des sédiments ainsi que les processus microbiens in situ sont rares et dispersées, en comparaison avec celles axées sur la composition de la communauté microbienne phylogénétique des sédiments pollués, et sur la biodégradation des polluants organiques en conditions expérimentales. Reste à savoir si la caractérisation de la communauté microbienne peut contribuer à une meilleure compréhension, prévision et modélisation des processus écosystémiques.
Les réponses microbiennes à la contamination des sédiments reposaient sur des adaptations directes aux conditions environnementales de toutes les principales caractéristiques structurelles et fonctionnelles, avec des répercussions potentielles sur l’efficacité du réseau trophique microbien dans le transfert d’énergie vers le réseau trophique benthique. Les modèles de composition de la MO des sédiments ont influencé les propriétés fonctionnelles des communautés benthiques. Les polluants organiques prioritaires, avec leurs effets toxiques prévisibles, pourraient jouer un rôle important dans le cycle C.
Élaborer et appliquer un système de classification pour classer les effets biologiques des perturbateurs endocriniens sur les sébastes mâles.Sebastiscus marmoratusdans la mer de Maowei, Chine
https://www.sciencedirect.com/science/article/abs/pii/S0025326X2031050X
Les perturbateurs endocriniens (EDC) présents dans les environnements marins sont devenus une préoccupation environnementale majeure. Néanmoins, les effets biologiques des perturbateurs endocriniens sur les organismes des milieux côtiers restent mal caractérisés. Dans cette étude, la biosurveillance des EDC chez le poisson mâle Sebastiscus marmoratus a été réalisée dans la mer de Maowei, en Chine. Les résultats ont montré que la concentration de 4-nonylphénol (4-NP) était inférieure à la limite de détection, que les concentrations de 4-tert-octylphénol (4-t-OP) et de bisphénol A (BPA) dans l’eau de mer étaient modérées par rapport à celles de l’eau de mer. d’autres régions du monde, et les sources possibles sont les rejets d’eaux usées municipales . Les analyses ANOVA imbriquées suggèrent des différences significatives dans les activités de l’aromatase cérébrale et l’expression de la vitellogénine plasmatique (VTG) entre la zone portuaire et la zone ostréicole . Un nouveau système expert sur les poissons (FES) a été développé pour évaluer les effets biologiques des perturbateurs endocriniens sur les poissons. Nos résultats montrent que le FES est un outil potentiel pour évaluer les effets biologiques des polluants marins.
Il existe une préoccupation constante concernant le rejet de composés perturbateurs endocriniens (EDC) dans le milieu aquatique en raison de leur potentiel à perturber le système endocrinien des espèces aquatiques. La principale source de perturbateurs endocriniens est d’origine humaine et leur présence dans le milieu aquatique est bien documentée. Les xénoestrogènes constituent une classe particulière d’EDC ayant la capacité d’imiter les œstrogènes naturels, et ils ont fait l’objet d’une attention particulière. Ces perturbateurs endocriniens comprennent certains pesticides ; les alkylphénols (AP); des plastifiants, tels que les phtalates et le bisphénol A (BPA) ; les congénères des polychlorobiphényles (PCB); les hydrocarbures aromatiques polycycliques (HAP) ; les métalloestrogènes, tels que le cadmium (Cd), le plomb (Pb), le calcium (Ca), le cobalt (Co), le cuivre (Cu), le nickel (Ni), le chrome (Cr), le mercure (Hg) et l’étain (Sn) ; et de nombreux autres produits chimiques industriels. Les perturbateurs endocriniens peuvent induire une série de changements biologiques chez les poissons, du niveau moléculaire jusqu’à l’ensemble de l’organisme, notamment des altérations des niveaux de transcription et de protéines, des changements dans les concentrations de stéroïdes sexuels en circulation, des changements histopathologiques dans les tissus cibles, un développement retardé ou altéré, un échec de reproduction et comportement anormal. Les perturbateurs endocriniens ont même été impliqués dans des effets au niveau des populations, notamment l’effondrement et le rétablissement des populations de poissons.
L’aromatase, une enzyme stéroïdogène codée par le gène CYP19, fait partie de la superfamille des enzymes du cytochrome P450. Comparés à la plupart des vertébrés, les poissons possèdent plusieurs isoformes de l’aromatase P450 (P450arom) structurellement et fonctionnellement distinctes, dérivées de locus génétiques distincts ( cyp19a et b ) et exprimées de manière différentielle dans le cerveau (P450aromB ≫ P450aromA) et les ovaires (P450aromA ≫ P450aromB) ; ces isoformes ont une expression constitutive (P450aromB ≫ P450aromA) et des programmes de développement différents. Il a été démontré que l’aromatisation de la testostérone (T) en œstrogènes localement dans le cerveau active les comportements sexuels masculins chez diverses espèces de vertébrés. Les effets in vivo de l’aromatisation de T sont cohérents avec la présence de motifs de liaison au récepteur des œstrogènes (ER) (éléments sensibles aux œstrogènes, ERE) dans le promoteur cyp19b , ce qui implique que le cyp19b est la cible principale d’un sous-ensemble d’EDC. P450aromA est principalement exprimé dans les ovaires et est impliqué dans la différenciation sexuelle et la croissance des ovocytes. Ainsi, sans connaissance préalable de sa biodisponibilité, de son accumulation, de son métabolisme ou de ses caractéristiques de liaison aux récepteurs, P450aromB est un biomarqueur pratique pour indiquer l’effet des EDC, ce qui constitue un point d’entrée pour découvrir les perturbations de la reproduction et du développement induites par les EDC.
Chez les vertébrés, la vitellogénine (VTG) est normalement synthétisée en réponse aux œstrogènes endogènes et est le principal précurseur des vitellines, qui fournissent les réserves énergétiques nécessaires au développement embryonnaire. Le VTG est généralement présent dans le plasma des poissons femelles. Les poissons mâles possèdent également le gène vtg , mais produisent peu d’œstrogènes en circulation et ne produisent normalement pas de VTG. Les mâles sont capables de synthétiser des quantités de VTG équivalentes à celles des organismes femelles matures lorsque les poissons sont exposés à des œstrogènes exogènes. De plus, il a été avancé que des teneurs anormales en VTG chez les poissons mâles et juvéniles, ainsi que des modifications de l’équilibre hormonal sexuel, sont des biomarqueurs fiables et complémentaires des effets des perturbateurs endocriniens dans des études en laboratoire et sur le terrain.
Dans l’ensemble, les résultats de la présente étude ont démontré des impacts variables des perturbateurs endocriniens sur la santé reproductive des poissons de la mer de Maowei. Il existe des différences significatives dans les activités de l’aromatase cérébrale et les niveaux d’expression plasmatique de VTG entre la zone portuaire et la zone ostréicole par analyses ANOVA imbriquées. Ces effets biologiques peuvent être liés aux différents produits chimiques possédant des propriétés perturbatrices endocriniennes, qui sont rejetés par les activités urbaines et industrielles, notamment dans les zones urbaines.
Effets écologiques des antibiotiques sur les écosystèmes naturels : une revue
https://www.sciencedirect.com/science/article/abs/pii/S0026265X17301108
Parmi les différents produits pharmaceutiques présents dans les écosystèmes du sol et de l’eau en tant que microcontaminants, une attention considérable a été accordée aux antibiotiques, car leur utilisation croissante et le développement conséquent de bactéries multirésistantes présentent de graves risques pour la santé humaine et vétérinaire. De plus, une fois introduits dans l’environnement, les antibiotiques peuvent affecter les communautés microbiennes naturelles. Ces derniers jouent un rôle clé dans les processus écologiques fondamentaux, notamment le maintien de la qualité des sols et de l’eau. En fait, ils sont impliqués dans le cycle biogéochimique et la dégradation des contaminants organiques grâce à leur vaste réservoir de diversité génétique et leur capacité métabolique. Lorsque des antibiotiques sont présents dans l’environnement, ils peuvent entraver la structure et le fonctionnement des communautés microbiennes de différentes manières et avoir des effets à la fois directs (à court terme) et indirects (à long terme) sur les communautés microbiennes. Les actions à court terme sont des actions bactéricides et bactériostatiques avec pour conséquence la disparition de certaines populations microbiennes et de leur fonctionnement écologique. L’impact indirect comprend le développement de bactéries résistantes aux antibiotiques et, dans certains cas, de souches bactériennes capables de les dégrader par des processus métaboliques ou co-métaboliques. La biodégradation permet d’éliminer complètement un composé toxique de l’environnement s’il est minéralisé.
Plusieurs facteurs peuvent influencer l’importance de ces effets directs et indirects, notamment la concentration de l’antibiotique, la durée d’exposition, l’écosystème récepteur (par exemple le sol ou l’eau) et la cooccurrence d’autres antibiotiques et/ou d’autres contaminants. Cette revue décrit l’état actuel des connaissances concernant les effets des antibiotiques sur les communautés microbiennes naturelles dans les écosystèmes du sol et de l’eau.
Les antibiotiques actuellement utilisés sont des molécules naturelles, synthétiques et semi-synthétiques. Les antibiotiques naturels sont produits par des bactéries et des champignons (par exemple la benzylpénicilline et la gentamicine) pour inhiber ou tuer d’autres micro-organismes concurrents (avec effet bactériostatique ou bactéricide). Les composés semi-synthétiques sont des antibiotiques naturels modifiés chimiquement par l’insertion d’un additif au sein de la formulation du médicament, ce qui améliore son efficacité (plus stable et moins biodégradable).
Les antibiotiques sont des molécules complexes qui peuvent avoir différents groupes fonctionnels au sein de leur structure chimique et peuvent être divisées en différentes catégories, en fonction de leur mécanisme d’action : l’inhibition de la synthèse de la paroi cellulaire, l’altération des membranes cellulaires, l’inhibition de la synthèse des protéines, la synthèse des inhibition des acides nucléiques et antagonisme métabolique ou anticoncurrentiel.
Après administration, ils ne sont que partiellement métabolisés et, par conséquent, une grande quantité est excrétée sous forme inchangée ou sous forme de métabolites actifs via l’urine et les fèces ; par conséquent, les antibiotiques humains atteignent les stations d’épuration des eaux usées (STEP). Les STEP conventionnelles ne sont pas spécifiquement conçues pour l’élimination des antibiotiques et, par conséquent, ces molécules sont libérées directement dans l’environnement récepteur. Bien que les STEP soient considérées comme la principale source d’antibiotiques et de gènes de résistance aux antibiotiques pour les eaux de surface, la législation actuelle au niveau européen ne contient pas d’exigence de concentration d’antibiotiques pour le rejet des STEP dans les eaux réceptrices. De nombreuses recherches ont été axées sur le développement de technologies innovantes pour l’élimination des antibiotiques dans les STEP. Ces traitements supplémentaires augmentent clairement les coûts de prise en charge et, pour cette raison, sont actuellement sous-utilisés. Les eaux usées des hôpitaux et de l’industrie pharmaceutique, l’élimination incontrôlée et illégale de médicaments et l’aquaculture peuvent également être une source importante de contamination aquatique. L’épandage de fumier et de boues sur le sol comme engrais, ainsi que l’irrigation avec de l’eau récupérée, peuvent contribuer à la dissémination des antibiotiques et des gènes de résistance aux antibiotiques dans le sol. Il a également été souligné qu’il existe une possibilité d’absorption d’antibiotiques par les plantes ou d’autres organismes à partir de sols fertilisés avec du fumier animal ou des eaux usées d’irrigation. Bien que les effets néfastes de l’ingestion d’antibiotiques présents dans les plantes ne soient pas bien connus et étudiés, ils pourraient provoquer des réactions allergiques ou toxiques et/ou améliorer la résistance aux antibiotiques chez l’homme. Enfin, à partir des sols superficiels, en fonction de leurs caractéristiques intrinsèques spécifiques, les antibiotiques peuvent être lessivés vers les couches plus profondes et les eaux souterraines. Dans cette revue, nous présentons des études récentes sur la réponse des communautés microbiennes naturelles aux antibiotiques en termes d’effets néfastes, de résistance et/ou de capacité de dégradation.
Pour minimiser la résistance aux antibiotiques, intensifiée par l’utilisation d’antibiotiques en médecine vétérinaire, l’UE a interdit depuis 2006 l’utilisation d’antibiotiques comme stimulateurs de croissance car l’administration à faibles doses (concentration minimale inhibitrice) pour moduler le métabolisme de la flore bactérienne commensale peut favoriser la propagation de ce phénomène. Malheureusement, dans d’autres pays (par exemple en Amérique, au Canada et en Asie), ils sont encore largement utilisés comme facteurs de croissance. La libération d’antibiotiques et de gènes de résistance dans les écosystèmes naturels est un événement récent en termes d’évolution. Leur impact sur les bactéries non ciblées et leurs fonctions écologiques associées suscite une préoccupation particulière. Ces polluants peuvent directement (effet bactéricide et bactériostatique avec disparition ou inhibition de certains groupes microbiens impliqués dans des fonctions clés de l’écosystème) ou indirectement (sélection de résistances, génération de variabilité génétique et phénotypique).
Risques environnementaux et toxicité des tensioactifs : aperçu des techniques d’analyse, d’évaluation et de remédiation
https://link.springer.com/article/10.1007/s11356-021-16483-w [41]
Les tensioactifs font partie des contaminants émergents les plus problématiques qui sont continuellement rejetés dans l’environnement par les usines de traitement des eaux usées (STEP). Les tensioactifs possèdent à la fois des propriétés hydrophiles et hydrophobes (queue d’hydrocarbure non polaire) et sont donc considérés comme des molécules amphipathiques. Les tensioactifs sont utilisés pour diverses applications domestiques et industrielles en raison de leurs propriétés physicochimiques uniques. Cependant, la persistance des produits transformés dans un environnement est très préoccupante pour la durabilité environnementale et la santé des écosystèmes. En perspective, les tensioactifs sont essentiels à la production industrielle de détergents, de textiles, de peintures, de polymères, de produits pharmaceutiques, de pesticides, de papier et de produits de soins personnels. Ces tensioactifs pourraient réduire la tension interfaciale et stabiliser les mousses et/ou les émulsions ; ils sont donc importants pour certaines opérations essentielles telles que la récupération du pétrole et l’exploitation minière. La taille du marché mondial des tensioactifs s’élève actuellement à environ 42,1 milliards de dollars américains et devrait atteindre 52,4 milliards de dollars d’ici 2025. Il y a plusieurs années, on estimait que les tensioactifs présents dans les eaux usées domestiques et industrielles se situaient respectivement entre 1 et 10 mg/L et 300 mg/L. Il est possible que les tensioactifs pénètrent dans l’eau potable par les stations d’épuration des eaux usées, posant ainsi un risque pour la santé humaine, animale et aquatique. De plus, le devenir et la structure chimique des surfactants, des systèmes environnementaux (aérobies ou anaérobies) et des accepteurs d’électrons contribuent principalement à la persistance, à la dégradabilité et aux effets écologiques des surfactants dans l’environnement.
Certains tensioactifs disponibles dans le commerce constituent une grave menace environnementale et publique pour les humains et les écosystèmes. Par exemple, les tensioactifs anioniques, principalement les alkylbenzènesulfonates linéaires (LAS), provoquent des impacts biochimiques, pathologiques, physiologiques et autres sur les écosystèmes aquatiques/terrestres. De plus, le LAS provoque des irritations cutanées et des problèmes respiratoires et réduit la résistance du biote aquatique au stress environnemental, à la reproduction et aux processus de croissance. Les tensioactifs augmentent généralement la solubilité des contaminants et facilitent ainsi l’eutrophisation. De plus, les propriétés hydrophobes croissantes des tensioactifs augmentent proportionnellement leur toxicité. Ces vastes impacts suscitent par conséquent des inquiétudes en matière de santé publique et d’environnement concernant la forte concentration de tensioactifs.
Classiquement, diverses techniques ont été déployées pour décontaminer les tensioactifs des eaux usées. Certaines de ces technologies d’assainissement comprennent des processus physico-chimiques, une filtration membranaire et une floculation-adsorption, une ultrafiltration, une électrocoagulation et une oxydation chimique et électro-oxydante. Cependant, lorsque les concentrations de tensioactifs dans les eaux usées sont élevées, presque toutes les techniques traditionnelles n’ont pas réussi à éliminer complètement la contamination par les tensioactifs. En général, le choix de technologies de traitement efficaces dépend en grande partie de la qualité de l’influent et de l’effluent. Les technologies de traitement physico-chimique possèdent d’excellentes efficacités d’élimination avec quelques défis notables. Par exemple, presque tous les traitements physiques et chimiques nécessitent des coûts d’investissement et des temps de rétention élevés et génèrent des boues. Cependant, le traitement physique nécessite moins d’intrants chimiques et opérationnels, tandis que le traitement chimique génère peu de boues. D’autre part, la facilité d’utilisation, le faible coût et l’efficacité d’élimination positionnent l’unité de traitement biologique et le système de boues activées cycliques comme une meilleure approche d’assainissement des tensioactifs. Cependant, le temps de rétention élevé, la formation de mousse, la mort de la biomasse et la génération massive de boues réduisent l’acceptation et l’applicabilité du traitement biologique. Des études antérieures ont prouvé que l’utilisation d’organismes thermophiles peut réduire les coûts opérationnels et la génération de boues et ainsi augmenter l’adoption du traitement biologique. L’adsorption est largement acceptée en raison de son excellente efficacité, de sa facilité d’utilisation et de modification des adsorbants, de sa rentabilité et de son respect de l’environnement. Auparavant, les nanotubes de carbone, la résine polymère, les cendres volantes-TiO 2 , les cendres volantes de charbon et les microsphères de chitosane réticulées par des acides aminés ont été utilisés pour éliminer les tensioactifs. Cependant, le charbon actif reste le candidat le plus approprié parmi de nombreux adsorbants développés pour éliminer les tensioactifs des eaux usées. Néanmoins, des efforts concertés sont nécessaires pour développer des adsorbants avancés dotés d’une efficacité exceptionnelle d’élimination des tensioactifs sans compromettre la sécurité environnementale et la santé publique.
Les tensioactifs peuvent pénétrer dans l’environnement à partir des effluents générés par les produits agrochimiques, les produits industriels et les activités domestiques et par conséquent provoquer une pollution environnementale. Par exemple, les biocides, les herbicides et les pesticides font partie des produits agrochimiques, tandis que les produits de soins personnels, les émulsifiants, les agents mouillants, les détergents et les revêtements ou adoucissants des tissus, du papier et des tapis sont des produits industriels notables qui contribuent continuellement à la protection de l’environnement grâce aux tensioactifs. pollution. De plus, la lessive, la désinfection et la fumigation sont d’importantes activités domestiques qui rejettent des tensioactifs dans l’environnement. Le devenir, la distribution et la persistance des tensioactifs dans l’environnement sont principalement influencés par la sorption et la bio/photodégradation. Ces processus dépendent principalement de divers facteurs environnementaux tels que le pH, la température et la salinité. Les stations d’épuration municipales reçoivent généralement une forte concentration de tensioactifs mais rejettent une faible concentration de tensioactifs et de leurs produits dégradés lors du traitement secondaire. La proportion de tensioactifs et de sous-produits post-traitement rejetés dépend davantage de l’efficacité des SMEEU.
Les produits transformés d’acides sulfophénylcarboxyliques, d’éthoxylates de nonylphénol, d’éthoxylates d’octylphénol et d’acides carboxyliques de nonylphénol ont été récemment signalés. Notamment, les éthoxylates de nonyl- ou d’octylphénol (NPEO) sont plus toxiques que leurs précurseurs, c’est-à-dire les alkylphénols (AP). Plusieurs études ont montré de graves impacts écologiques et sanitaires des tensioactifs toxiques sur les humains, d’autres vertébrés, la faune du sol, les micro-organismes, les crustacés et les plantes terrestres. Les critères critiques pour évaluer les risques environnementaux associés aux tensioactifs disponibles dans le commerce sont la photo ou la biodégradabilité, le comportement toxique et l’efficacité de sorption. Par exemple, la formation de mousse et la sédimentation des tensioactifs détériorent la qualité de l’eau en réduisant le transfert d’oxygène de l’air ou de l’eau et la capacité autonettoyante des rivières, à des degrés différents selon les classes de tensioactifs. De plus, les tensioactifs améliorent la solubilité des polluants organiques persistants (POP) dans la phase aqueuse, et les produits d’aérosols et de tensioactifs qui en résultent ont un impact significatif sur l’atmosphère et le climat. Les changements dans les activités physiologiques et biochimiques des organismes aquatiques par le LAS retardent leur métabolisme et leur croissance, endommagent la membrane cellulaire et provoquent la rupture du complexe protéique de la chlorophylle.
Selon le règlement REACH (enregistrement, évaluation, autorisation et restriction des produits chimiques), l’écotoxicité des tensioactifs peut être évaluée à l’aide de micro-biotests (QSAR, ECOSAR). Les procédures utilisent des tests tels que Daphtoxkit F, Daphnia IQ Test, Rotokit F/M, Ceriodaphtoxit K, Ostracode toxkit, Algal toxkit F et Microtoxkit. Les études antérieures ont montré que la toxicité des tensioactifs pouvait être influencée par les propriétés physicochimiques de l’eau (c’est-à-dire le pH, l’OD, les matières en suspension), les tensioactifs (c’est-à-dire le type, la concentration et la capacité d’absorption des tensioactifs) et des facteurs biotiques (l’âge et le type d’espèces). , sensibilité entre les espèces et leur acclimatation. La concentration médiane efficace (EC50) et la concentration médiane d’inhibition de la croissance (IC50) déterminent respectivement le risque et la toxicité des tensioactifs sur les animaux, les plantes et les micro-organismes : la CE50 à 96 h du chlorure d’acétyltriméthylammonium sur la microalgue verte Chlorella vulgaris est de 145 ± 13,35 μg/L, tandis que la CE50 du benzalkonium sur les invertébrés est de 5,90 μg/L. De plus, la CI50 du chlorure de benzyldiméthyldodécylammonium pour les bactéries est de 170,0 μg/L. Les tensioactifs peuvent retarder la croissance microbienne et augmenter les mutations microbiennes et la mortalité. Par exemple, les éthoxylates de nonylphénol (NPEO) peuvent dissocier la production d’énergie, ce qui peut par conséquent perturber les processus de croissance microbienne et de nitrification. De plus, les tensioactifs anioniques perturbent les structures internes et les fonctions microbiennes telles que la résistance environnementale, la croissance, le stress compétitif et la reproduction. L’absorption de tensioactifs par un micro-organisme peut dépolariser la membrane cellulaire microbienne et diminuer l’absorption des nutriments, l’acceptation de l’oxygène et la libération des métabolites toxiques. Certains tensioactifs ont de graves conséquences sur la santé humaine en raison de l’ingestion ou de la consommation d’aliments contaminés. Par exemple, les tensioactifs réagissent avec les protéines existantes dans le foie et le sérum, provoquant ainsi des effets métaboliques à long terme et une perturbation du système endocrinien humain. De même, il a été rapporté que certains tensioactifs provoquent des brûlures ou des irritations de la peau humaine ainsi que des problèmes oculaires et respiratoires. Les éthoxylates et carboxylates d’alkylphénol retardent la pénétration souterraine des composés pharmaceutiques. Les éthoxylates de nonylphénol ont des effets œstrogéniques sur les amphibiens, les mammifères et les Poissons. Certains tensioactifs tels que le LAS endommagent la membrane des cellules racinaires et modifient la structure de la membrane. Par conséquent, cela a altéré la transpiration et la translocation des nutriments essentiels et de l’eau.
Le rejet excessif de tensioactifs dans l’environnement met en danger la santé humaine et la sécurité écologique. Par conséquent, plusieurs techniques d’évaluation sont utilisées pour séparer, identifier et quantifier les tensioactifs dans une matrice environnementale. Cette section se concentre sur diverses techniques d’évaluation grâce auxquelles différents tensioactifs présents dans des échantillons environnementaux peuvent être quantifiés. L’intégrité et l’efficacité des techniques d’évaluation dépendent principalement du niveau de contamination environnementale et de l’efficacité des technologies d’assainissement. La présence de tensioactifs dans les environnements aquatiques et terrestres a des effets néfastes sur les micro-organismes, les plantes, les animaux et les humains. L’immense utilisation des tensioactifs, leurs risques pour la santé et les politiques réglementaires strictes concernant leur limite autorisée nécessitent leur contrôle dans l’environnement. Cette limite admissible a fait l’objet de débats entre plusieurs chercheurs. Par conséquent, plusieurs approches d’assainissement ont été recommandées pour nettoyer les tensioactifs présents dans l’environnement. Au cours des dernières décennies, la bioremédiation a souvent été utilisée pour éliminer les tensioactifs de la matrice environnementale. Récemment, les méthodes d’assainissement disponibles englobent les techniques physiques, chimiques, biologiques et membranaires. Les approches d’assainissement biologique et physique, y compris la décantation par gravité, ne peuvent pas éliminer complètement les tensioactifs anioniques et les matières organiques lorsque la concentration de polluants est élevée. La méthode d’assainissement appropriée est influencée par de nombreux facteurs tels que l’efficacité du traitement, la durée de fonctionnement, les caractéristiques des effluents et des affluents et la consommation d’énergie.
La présence de tensioactifs dans les environnements aquatiques et terrestres a des effets néfastes sur les micro-organismes, les plantes, les animaux et les humains. L’immense utilisation des tensioactifs, leurs risques pour la santé et les politiques réglementaires strictes concernant leur limite autorisée nécessitent leur contrôle dans l’environnement. Cette limite admissible a fait l’objet de débats entre plusieurs chercheurs. Par conséquent, plusieurs approches d’assainissement ont été recommandées pour nettoyer les tensioactifs présents dans l’environnement. Au cours des dernières décennies, la bioremédiation a souvent été utilisée pour éliminer les tensioactifs de la matrice environnementale. Récemment, les méthodes d’assainissement disponibles englobent les techniques physiques, chimiques, biologiques et membranaires. Les approches d’assainissement biologique et physique, y compris la décantation par gravité, ne peuvent pas éliminer complètement les tensioactifs anioniques et les matières organiques lorsque la concentration de polluants est élevée. La méthode d’assainissement appropriée est influencée par de nombreux facteurs tels que l’efficacité du traitement, la durée de fonctionnement, les caractéristiques des effluents et des affluents et la consommation d’énergie.
Cet examen complet a révélé les énormes implications écotoxicologiques et sur la santé humaine des tensioactifs dans l’environnement. La non-dégradabilité, la persistance et la toxicité de certains tensioactifs et de leurs produits transformés ont des conséquences environnementales extrêmes. Ainsi, l’équilibre et la durabilité des écosystèmes naturels pourraient être gérés grâce à l’évaluation, à l’analyse pratique et au traitement des tensioactifs. Par conséquent, plusieurs technologies d’analyse, d’évaluation et d’assainissement sont utilisées pour identifier, quantifier et nettoyer efficacement les tensioactifs des matrices environnementales. Sur la base de la littérature disponible, ces travaux ont montré que les techniques chromatographiques, en particulier la chromatographie fluide supercritique et quadripolaire à temps de vol MS (Q-ToF-MS), offrent une meilleure analyse des tensioactifs jusqu’à des concentrations en traces dans les matrices environnementales. Compte tenu des atouts et des défis des diverses méthodes d’assainissement disponibles pour les tensioactifs, le fractionnement des mousses, les réacteurs thermiques à membrane aérobie, les adsorbants avancés et les processus aérobies électrochimiques avancés sont des méthodes efficaces. Les recherches futures devraient se concentrer sur le développement d’adsorbants bon marché à partir de déchets. Cependant, des recherches approfondies sur leur récupération, régénération, réutilisation et valeurs technico-économiques devraient être envisagées. En outre, il est pertinent d’intégrer ou de coupler les technologies avancées d’assainissement actuelles aux usines de traitement des eaux usées (STEP) existantes pour une élimination efficace des tensioactifs des affluents. L’oxydation par faisceau d’électrons génère des radicaux puissants capables de minéraliser complètement les tensioactifs. Cependant, ses mécanismes, sa cinétique et ses coûts n’ont pas été suffisamment étudiés et documentés. Enfin, cette revue vise à favoriser une bonne compréhension des analyses et évaluations réalisables ainsi que des approches d’assainissement réalisables pour aider les chercheurs, les décideurs politiques et les organismes de réglementation dans leurs efforts visant à minimiser l’exposition environnementale aux tensioactifs toxiques.
Différentes conditions d’optimisation requises pour améliorer la biodégradation des tensioactifs linéaires alkylbenzosulfonate et dodécylsulfate de sodium par un nouveau consortium d’ Acinetobacter calcoaceticus et Pantoea agglomerans
https://www.sciencedirect.com/science/article/abs/pii/S0141022907001184?via%3Dihub [42}
Les tensioactifs anioniques impliquent un groupe de composés xénobiotiques qui possèdent des groupes sulfonés ou ester sulfate qui sont des ingrédients largement utilisés dans plusieurs utilitaires industriels. En raison de leur grande consommation dans le monde, les tensioactifs anioniques ont le potentiel d’être largement expulsés dans les environnements aquatiques et terrestres. L’alkylbenzène sulfoné linéaire (LAS) est l’un des principaux tensioactifs anioniques xénobiotiques qui ont des applications industrielles à grande échelle et donc une large diffusion dans l’environnement. Ce tensioactif est en grande partie rejeté lors du traitement des eaux usées, mais une partie des eaux usées peut rejeter du LAS directement dans l’environnement. Des effets toxiques possibles du LAS ont été rapportés dans des sols agricoles recevant des charges élevées de ce tensioactif provenant de boues d’épuration recyclées ou d’eau d’irrigation contaminée. Apparemment, la voie de biodégradation microbienne représente le principal mécanisme d’élimination des LAS provenant d’un environnement pollué. Structurellement, le LAS commercial contient une chaîne alkyle linéaire, un cycle benzénique et un groupe sulfoné. Un aperçu de la voie de biodégradation du LAS a été élaboré, qui commence par une oxygénation à l’extrémité terminale de la chaîne alkyle suivie d’une β-oxydation et d’une ouverture du cycle ainsi que d’une conversion du groupe sulfoné en sulfate inorganique. En raison des besoins stricts en oxygène au début de cette voie, la biodégradation du LAS est généralement plus favorisée dans des conditions aérobies qu’anaérobies. Des expériences avec des boues d’eau de mer ont montré que les communautés bactériennes dégradent le LAS avec une plus grande efficacité que les souches isolées et donc, un consortium de plusieurs souches bactériennes est normalement nécessaire pour réaliser la minéralisation de ce tensioactif en conditions aérobies. Les isolats bactériens uniques ont généralement une capacité limitée à dégrader la chaîne alkyle et n’ont pas non plus la capacité de cliver le cycle aromatique sulfoné du LAS.
Les composés de sulfate d’alkyle aliphatique communément représentés par le dodécylsulfate de sodium (SDS) sont d’autres tensioactifs anioniques qui peuvent facilement se retrouver dans l’environnement par les eaux usées domestiques et industrielles. Ils se caractérisent par leur structure linéaire d’ester sulfate à longue chaîne qui peut être dégradée par plusieurs souches bactériennes par désulfatation et assimilation ultérieure. Certaines souches d’Acinetobacter sont connues pour être impliquées dans la biodégradation de différents polluants tels que le biphényle ou le chlorobiphényle, l’aniline, le phénol, le benzoate, le pétrole brut et l’acétonitrile. Acinetobacter calcoaceticus , une souche anaérobie facultative capable de dégrader davantage les alcanes que les fractions aromatiques du pétrole brut. Il a bien poussé sur les n -alcanes jusqu’au C33 ainsi que sur les alcanes à chaîne ramifiée, mais n’a pas réussi à croître sur les cycloalcanes. La Pantoea agglomerans est une autre souche anaérobie facultative, associée à la biodégradation du kérosène, du toluène et de la vaseline.
Bien que des travaux considérables aient été menés sur la biodégradation individuelle des tensioactifs anioniques aromatiques LAS ou SDS aliphatiques, peu d’attention a été accordée à la comparaison des conditions de biodégradation de ces deux composés structurellement différents. Il n’existait pas non plus de documentation sur un système bactérien unique capable de biodégrader les deux tensioactifs. Le présent travail a été axé sur ces aspects particuliers de la biodégradation du LAS et du SDS en utilisant un nouveau consortium de cultures mixtes contenant A. calcoaceticus et P. agglomerans .
Les deux tensioactifs, à des concentrations plus élevées, sont devenus toxiques pour les bactéries de co-culture mixte, affectant particulièrement la souche A. calcoaceticus du consortium. La supplémentation de l’un ou l’autre tensioactif comme seule source de carbone à la culture mixte poussant dans un milieu minimal a produit respectivement 10 % de dégradation du SDS et 60 % de dégradation du LAS. Ces données sont discutées en termes de différences entre l’induction enzymatique de la biodégradation bactérienne LAS et SDS.
Impacts de la pollution plastique sur la biogéochimie du carbone marin
https://www.sciencedirect.com/science/article/abs/pii/S0269749120362862
L’un des défis majeurs dans la compréhension de la dynamique de la santé et du fonctionnement des océans est l’impact potentiel de la présence croissante de plastique. Outre leurs effets vérifiés et macroscopiques sur la faune et les habitats marins, les micro et macroplastiques offrent des sites potentiels pour l’activité microbienne et le lessivage chimique . La plupart des plastiques marins se trouvent initialement dans les mètres supérieurs de la colonne d’eau, où les processus biogéochimiques fondamentaux déterminent la productivité marine et la dynamique du réseau trophique. Cependant, des découvertes récentes montrent un continuum d’effets potentiels de ces nouveaux composants marins sur le carbone, les nutriments et les processus microbiens. Dans la présente analyse, nous développons un terrain d’entente entre ces études et identifions les lacunes dans les connaissances sur lesquelles de nouveaux efforts de recherche devraient être concentrés, afin de mieux déterminer les rétroactions potentielles des plastiques sur la biogéochimie du carbone d’un océan en évolution. Une accumulation de plastique à la surface de la mer, un environnement déjà enrichi en matière organique dérivée de l’activité microbienne dans la zone euphotique, favorise en outre la croissance microbienne et la libération de polymères organiques complexes à l’interface air-mer. Ce processus implique 1) une pénétration réduite du rayonnement photosynthétiquement actif (PAR) pour la croissance du phytoplancton et la photosynthèse, 2) une production altérée d’oxygène et de dioxyde de carbone liée à des processus de production/dégradation altérés, et 3) une barrière plus épaisse pour l’air. échange marin de gaz. De plus, le carbone organique dissous (COD) extrait des plastiques augmente l’activité bactérienne hétérotrophe et le remaniement de la matière organique dissoute (MOD), avec pour conséquence une augmentation de la respiration bactérienne et de la consommation d’oxygène. À des profondeurs plus élevées, l’agrégation de la matière organique autour des particules en suspension, comme le plastique, est facilitée par l’interaction du phytoplancton et des bactéries. 4) Le biofilm qui intègre la particule de plastique modifie sa flottabilité et facilite son flux descendant, tout en transférant le carbone organique particulaire (POC) vers l’intérieur de l’océan et éventuellement vers le fond marin. 5) Le biofilm entourant la particule de plastique peut incorporer des nutriments supplémentaires tout au long de son flux descendant, favorisant ainsi une activité microbienne supplémentaire et la libération de composés organiques plus complexes, comme la DOM réfractaire (RDOM), alimentant ainsi un réservoir de carbone réfractaire à long terme en profondeur. Le plastique peut affecter les organismes marins, du micro au macro, à travers de multiples processus qui impliquent l’ingestion, l’enchevêtrement, la destruction de l’habitat, le transfert de composés toxiques, soit par la libération de composants du plastique lui-même, soit par la concentration et la libération de polluants présents dans l’eau de mer environnante. Des recherches récentes montrent également que les particules de plastique peuvent transporter des espèces pathogènes, conduisant au développement possible d’une résistance aux antibiotiques avec d’autres effets délétères sur la santé.
L’impact des interactions microplastiques-microbes sur la santé animale et les cycles biogéochimiques
https://www.sciencedirect.com/science/article/abs/pii/S0048969721007658
La pollution par les microplastiques (MP) a attiré l’attention mondiale en raison de l’utilisation intensive de produits en plastique. La surface hydrophobe du MP fournit un habitat à plusieurs micro-organismes. Bien qu’il existe plusieurs études sur l’impact des particules de plastique sur les communautés microbiennes cycles biogéochimiques, rares sont les revues qui résument systématiquement l’interaction entre les MP et les microbes et leurs effets sur la santé humaine et la circulation biochimique. Les discussions dans cette revue se dérouleront sur les sujets suivants : (1) les députés provoquent la colonisation, la génération de biofilms et le transfert de microbes environnementaux ; (2) les communautés microbiennes peuvent provoquer des altérations morphologiques et une biodégradation des MP ; (3) les combinaisons MP-microbes peuvent induire une altération de la flore intestinale et mettre en danger la santé animale ; (4) l’affecté par les interactions MP-microbe. Cette revue mettra en évidence les interactions étroites entre les MP et les micro-organismes et fournira des suggestions pour de futures études.
plusieurs rapports ont montré que les MP constituaient des dangers potentiels pour le biote du milieu environnant : les déchets plastiques réduisaient le taux de croissance des vers de terre, entraînant ainsi une perte de poids, ce qui peut être particulièrement nocif pour leur capacité de reproduction. le plus important, les communautés microbiennes affectées par les débris plastiques méritent une attention particulière. L’abondance des micro-organismes joue un rôle important dans les systèmes écologiques, notamment le métabolisme des substances, la formation de produits et le cycle trophique. Les environnements complexes contiennent des structures et des catégories distinctes de microbes, les communautés microbiennes de l’environnement étant dynamiques. Les communautés microbiennes sont capables de réagir et de s’adapter rapidement aux changements de leur environnement causés par le stress anthropique ou le changement climatique. Dans l’écosystème terrestre, les MP pouvaient modifier la composition des communautés microbiennes en fonction des propriétés physiques du sol. Par exemple, les phylums de Bacteroidetes, Proteobacteria et Gemmatimonadetes ont été enrichis dans un sol modifié au polyéthylène, ce qui peut contribuer aux changements potentiels dans la teneur en DOM (matière organique dissoute), la densité apparente et l’humidité du sol. Dans le milieu aquatique, les plastiques favorisent la colonisation par des communautés microbiennes, notamment des bactéries pathogènes. L’invasion de bactéries pathogènes peut perturber les communautés bactériennes intestinales normales et diminuer la capacité de défense des organismes. Environ la moitié des 20 genres les plus abondants adhérant au PE étaient des agents pathogènes, illustrant que les plastiques agissaient comme vecteurs de transfert de micro-organismes nocifs, posant une menace potentielle pour la santé humaine.
Il existe actuellement plusieurs rapports sur les menaces posées par les plastiques, qui ont donné lieu à des variations de biocénose. Une grande quantité de nutriments adhère en peu de temps aux surfaces plastiques de l’eau de mer, ce qui attire la colonisation microbienne pour utiliser les substances nutritives ont été les premiers à définir cette communauté complexe comme « Plastisphère » et ont suggéré qu’il existait des variations dans les microbes trouvés à la surface des débris et dans leurs environs. Les plastiques peuvent fournir un habitat unique aux micro-organismes, qui utilisent davantage les nutriments environnants pour reproduire des progénitures et biodégrader les morceaux de plastique. Cependant, aucune analyse systématique n’a été réalisée sur les relations entre les MP et les micro-organismes, ni sur leurs impacts potentiels sur la santé animale et la circulation biochimique.
(1) les MP provoquent la colonisation bactérienne, génèrent des biofilms et accélèrent le transfert ; (2) les communautés microbiennes induisent des altérations physiques et la biodégradation des plastiques ; (3) les députés modifient la microflore intestinale et menacent la santé animale ; (4) les cycles du carbone et de l’azote affectés par les interactions MP-microbe. Cette revue mettra en évidence les relations entre les microplastiques et les communautés microbiennes, ainsi que les impacts sur la santé animale et la circulation biochimique. Le cycle biogéochimique fait référence au mouvement cyclique d’éléments chimiques essentiels entre les organismes et l’environnement extérieur, un processus qui contribue à la synthèse et à la dégradation de la matière organique. Dans l’écosystème naturel, l’utilisation de produits en plastique a considérablement amélioré l’interaction entre les microbes et les MP. Il est donc possible que les plastiques et leurs intermédiaires de dégradation fournissent des sources de carbone,
Les débris de plastique comme source mobile d’additifs chimiques dans les environnements marins : preuves in situ
https://www.sciencedirect.com/science/article/abs/pii/S0048969722059927
Divers additifs sont délibérément ajoutés aux plastiques à des fractions volumiques élevées (par exemple, le chlore polyvinylique contient jusqu’à 70 % de phtalates en poids) pour obtenir certaines propriétés souhaitables, et certains de ces additifs sont répertoriés comme perturbateurs endocriniens. produits chimiques (par exemple, les oligomères de styrène et les phtalates) ou les polluants organiques persistants (par exemple, les éthers diphényliques polybromés et les composés perfluorés ). Malgré l’opinion de plus en plus répandue selon laquelle l’ingestion de microplastiques n’est pas censée augmenter la bioaccumulation de produits chimiques dans les organismes marins, la lixiviation d’additifs chimiques à partir de gros débris de plastique peut augmenter les niveaux de produits chimiques dans les organismes marins. l’environnement et l’accumulation de ces produits chimiques dans les réseaux alimentaires, y compris chez l’homme.
Cycle du soufre
Le cycle du soufre est un cycle biogéochimique qui implique le mouvement du soufre entre les roches, les cours d’eau et les organismes vivants. Le soufre est l’un des éléments les plus répandus sur Terre. Ce non-métal est jaune, cassant, inodore et insipide. Le soufre est présent dans tout de protéines les types. La méthionine, la cystine et la cystéine contiennent du soufre acides aminés qui sont directement absorbés par les plantes. La libération de soufre dans l’atmosphère est causée par la combustion de combustibles fossiles, l’activité volcanique et la dégradation des molécules organiques. Le soufre est stocké dans les roches souterraines et les minéraux terrestres. Il est libéré à la suite des précipitations, de l’altération des roches et des évents géothermiques. Les composés soufrés peuvent être utilisés comme oxydants ou réducteurs dans la respiration microbienne. Le cycle global du soufre implique le changement d’espèces de soufre via plusieurs états d’oxydation, qui jouent un rôle important dans les processus géologiques et biologiques. Le cycle du soufre est décrit comme suit :Par l’altération des roches, le soufre est libéré.
Lorsque le soufre entre en contact avec l’air, des sulfates se forment Les plantes et les micro-organismes absorbent les sulfates et les transforment en composés organiques. La forme organique du soufre est ensuite ingérée par les animaux via leur nourriture, transférant ainsi le soufre dans le chaîne alimentaire. Lorsque les animaux meurent, une partie du soufre est libérée par décomposition et une partie pénètre dans les tissus des micro-organismes. Il existe diverses sources naturelles telles que les éruptions volcaniques, l’évaporation de l’eau et la dégradation des matières organiques. déchets dans les zones humides, qui libèrent du soufre directement dans l’atmosphère. Ce soufre tombe sur terre avec les précipitations.
- Les plantes et les micro-organismes absorbent les sulfates et les transforment en composés organiques.
- La forme organique du soufre est ensuite ingérée par les animaux via leur nourriture, transférant ainsi le soufre dans le chaîne alimentaire.
- Lorsque les animaux meurent, une partie du soufre est libérée par décomposition et une partie pénètre dans les tissus des micro-organismes.
- Il existe diverses sources naturelles telles que les éruptions volcaniques, l’évaporation de l’eau et la dégradation des matières organiques. déchets dans les zones humides, qui libèrent du soufre directement dans l’atmosphère. Ce soufre tombe sur terre avec les précipitations.
- La réduction assimilatrice de sulfate fait référence au processus par lequel les plantes et les micro-organismes réduisent le sulfate, l’espèce complètement oxydée, pour une utilisation dans la production d’acides aminés et de protéines.
- Lorsque le sulfate se diffuse dans des environnements anoxiques, la réduction du sulfate dissimilatoire microbienne devient possible.
- Au cours de respiration anaérobie, le sulfate sert d’accepteur d’électrons terminal pour un certain nombre de micro-organismes, notamment des protéobactéries telles que Desulfovibrio et Desulfonema spp. et archaea appartenant à la genre Archéoglobe.
- Cela conduit à l’accumulation de sulfure. Le sulfure entièrement réduit peut ensuite servir de source d’électrons pour les micro-organismes photosynthétiques anoxygéniques et les chimiolithoautotrophes, tels que les membres de la famille des Chlorobi. phylum et le genre Thiobacillus, respectivement.
- Ces micro-organismes convertissent le sulfure en soufre et en sulfate. D’autres bactéries ont été découvertes pour réduire le soufre élémentaire (S0) par des moyens dissimilatoires.
- Dans les sédiments hypersalins, ces organismes comprennent Desulfuromonas, des archées thermophiles et des cyanobactéries. Le sulfite (SO32) est un intermédiaire essentiel supplémentaire qui peut être converti en sulfure par un large éventail de microbes, y compris des représentants des genres Alteromonas, Clostridium, Desulfovibrio et Desulfotomaculum.
- Le diméthylsulfoniopropionate (DMSP) est un produit chimique soufré crucial molécule. produit comme soluté compatible par marine phytoplancton. Le bactérioplancton (bactéries flottantes) utilise le DMSP comme source de carbone et de soufre lorsque ces cellules meurent et le libèrent.
- Il est transformé en sulfure de diméthyle (DMS) puis rejeté dans l’atmosphère.
- Là, le DMS est rapidement transformé en un certain nombre de composés soufrés qui servent de noyaux pour la production de gouttelettes d’eau et contribuent à la formation de nuages.
- Il est théorisé que l’amélioration de la production de DMS pourrait compenser les conséquences du changement climatique mondial car les nuages aident à garder la surface de la Terre fraîche.
Cycle du soufre gazeux
- Par l’émission bactérienne (H2S), la combustion de combustibles fossiles (SO2), les sels marins soufflés par le vent (SO 2-4) et les émissions volcaniques, le soufre atteint l’atmosphère (H2S, SO2, SO2-4).
- La majorité du soufre existant sous forme de SO2 ou de H2S est transformée en SO3, qui se dissout dans les gouttelettes d’eau pour générer de l’acide sulfurique.
- En raison de l’utilisation de combustibles fossiles, le cycle du soufre est surchargé.
- Par conséquent, le SO2 rejeté dans l’atmosphère constitue une part importante du transport total du soufre à l’échelle mondiale. Ce niveau plus élevé de soufre est converti en acide sulfurique dans l’eau de pluie, ce qui entraîne des répercussions écologiques négatives.
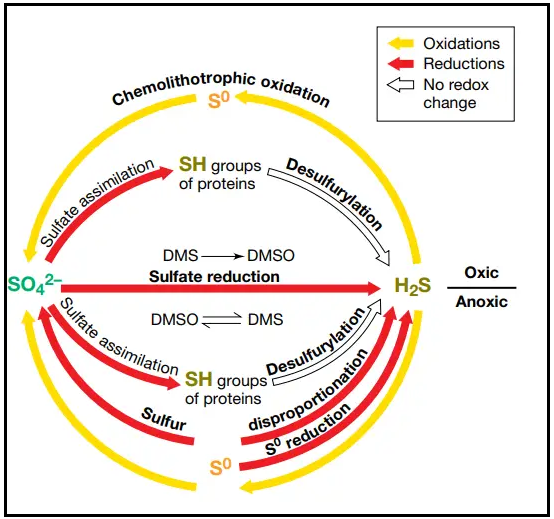
Cycle du soufre sédimentaire
- Dans la phase sédimentaire, l’altération et la dégradation des dépôts inorganiques et organiques libèrent du soufre.
- L’ion SO2-4 transporte le soufre vers les habitats terrestres et aquatiques. Après avoir été absorbé par les plantes et les microbes du sol, l’ion sulfate est réduit et finalement intégré sous forme de groupe sulfhydryle (-SH) dans protéines. DONC
- Les bactéries Desulfovibrio sur le fond de l’océan convertissent certains sulfates directement en sulfures, H2S ou S élémentaire dans des circonstances anaérobies.
- Cet H2S s’échappe dans l’atmosphère et reconstitue le soufre perdu par les précipitations.
- Les bactéries du genre Thiobacillus oxydent H2S avec O2 pour produire des sulfates. Le soufre en excès se mélange à l’eau pour produire des pluies acides.
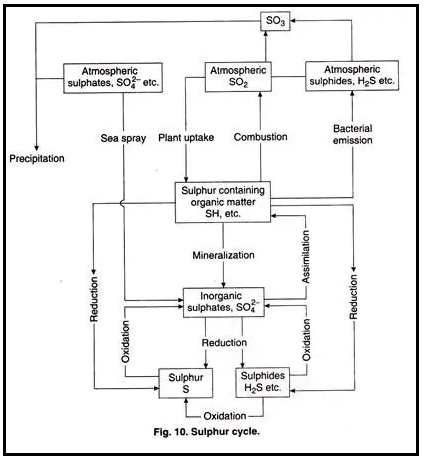
Décomposition des composés organiques
- La dégradation des protéines génère des acides aminés soufrés.
- Les bactéries Desulfotomaculum transforment les sulfates en sulfure d’hydrogène (H2S).
Oxydation du sulfure d’hydrogène en soufre élémentaire
- Le sulfure d’hydrogène s’oxyde pour donner du soufre élémentaire.
- Le processus d’oxydation est initié par certains bactéries photosynthétiques des familles Chlorobiaceae et Chromatiaceae.
Oxydation du soufre élémentaire
- Le soufre élémentaire présent dans le sol ne peut pas être directement utilisé par les plantes.
- Par conséquent, les bactéries chimiolithotrophes le transforment en sulfates.
Réduction des sulfates
- Desulfovibrio desulfuricans convertit les sulfates en sulfure d’hydrogène.
- Cela se déroule en deux étapes :
- Initialement, l’ATP est utilisé pour convertir les sulfates en sulfites.
- La conversion du sulfite en hydrogène sulfuré est la seconde étape.
Impact des activités humaines sur le cycle du soufre
- Le cycle global du soufre est fortement influencé par l’activité humaine.
- En raison de la combustion du charbon, du gaz naturel et des combustibles fossiles, la quantité de soufre dans l’atmosphère et l’océan a considérablement augmenté.
- De plus, le puits de roche sédimentaire a été épuisé.
- Sans l’influence de l’homme, le soufre resterait retenu dans les roches pendant des millions d’années jusqu’à ce qu’il soit soulevé par l’activité tectonique, puis libéré par les intempéries et l’érosion.
- Cependant, il est actuellement fouillé et brûlé à un rythme alarmant.
- Les dépôts de sulfates ont été multipliés par 30 dans les régions les plus polluées de la planète.
- Depuis le début du XXe siècle, le charbon et le pétrole ont été extraits à un rythme si rapide que le flux mondial de soufre a doublé.
Le diméthylsulfoniopropionate DMSP est un précurseur du dimétbylsulfure. DMSP est l’abréviation de diméthylsulfoniopropionate. Le diméthylsulfoniopropionate DMSP est un composé organosulfure répondant à la formule (CH3)2S + CH2CH2COO-.
Ce métabolite zwitterionique peut être trouvé dans le phytoplancton marin, les algues et certaines espèces de plantes vasculaires terrestres et aquatiques. Il fonctionne comme un osmolyte et a plusieurs autres rôles physiologiques et environnementaux qui ont également été identifiés. DMSP a d’abord été identifié dans l’algue rouge marine Polysiphonia fastigiata.
Chez les plantes supérieures, le DMSP est biosynthétisé à partir de la S-méthylméthionine. Deux intermédiaires dans cette conversion sont la diméthylsulfoniumpropylamine et le diméthylsulfoniumpropionaldéhyde. Chez les algues, cependant, la biosynthèse commence par l’élimination du groupe amino de la méthionine, plutôt que de la S-méthylméthionine.
DMSP est décomposé par des microbes marins pour former deux principaux produits volatils en soufre, chacun avec des effets distincts sur l’environnement. L’un de ses produits est le méthanethiol (CH3SH), qui est assimilé par les bactéries dans le soufre des protéines. Un autre produit de panne volatile est le sulfure de diméthyle (CH3SCH3, DMS). Il existe des preuves que le DMS dans l’eau de mer peut être produit par clivage du DMSP dissous (extracellulaire) par l’enzyme DMSP- lyase, bien que de nombreuses espèces de bactéries non marines convertissent le méthanethiol en DMS.
DMS est également pris en charge par les bactéries marines, mais pas aussi rapidement que le méthanethiol. Bien que DMS soit habituellement inférieur à 25 % des produits volatils de DMSP, la haute réactivité du méthanethiol rend les concentrations de DMS à l’état stationnaire dans l’eau de mer environ 10 fois celles du méthanethiol (~3 nM contre ~0,3 nM). Curieusement, il n’y a jamais eu de corrélations publiées entre les concentrations de DMS et de méthanethiol. Ceci est probablement dû à l’absorption abiotique et microbienne non linéaire du méthanethiol dans l’eau de mer et à la faible réactivité relativement faible du DMS. Cependant, une partie significative de DMS dans l’eau de mer est oxydée en diméthylsulfoxyde (DMSO).
En ce qui concerne le climat mondial, on pense que le DMS joue un rôle dans la protection de la Terre en réduisant la quantité de rayonnement solaire qui atteint la surface de la Terre. Cela intervient par la dégradation du DMS dans l’atmosphère en composés hygroscopiques qui condensent la vapeur d’eau conduisant à la formation de nuages.
DMSP a également été impliqué pour influencer les caractéristiques de goût et d’odeur de divers produits. Par exemple, bien que le DMSP soit inodore et insipide, il s’accumule à des niveaux élevés chez certains herbivores marins et notamment chez les filtreurs (bivalves). L’augmentation des taux de croissance, de la vigueur et de la résistance au stress chez les animaux cultivés dans de tels régimes ont été signalés. DMS, est responsable de l’hydrofuge, des goûts et des odeurs qui se développent avec certains produits de la mer en raison de l’action de la DMSP-lyase bactérienne qui cogénère l’acrylate.
Le diméthylsulfoniopropionate (DMSP) est un composé organosulfuré de formule chimique (CH3)2S+CH2CH2COO−. Ce zwitterion est un métabolite du plancton, des algues et de certaines espèces de plantes vasculaires terrestres et aquatiques. Il assure diverses fonctions physiologiques et joue notamment le rôle d’osmolyte2.
Le DMSP est synthétisé par les plantes vasculaires à partir de la S-méthylméthionine par l’intermédiaire de la diméthylsulfoniumpropylamine et du diméthylsulfoniumpropionaldéhyde3. Chez les algues, cependant, sa biosynthèse commence par l’élimination du groupe amine de la méthionine.
Il est dégradé par les microorganismes marins pour former deux composés soufrés volatils principaux ayant chacun des effets distincts sur l’environnement. Il se forme essentiellement du méthanethiol CH3SH, assimilé par les bactéries en soufre protéique, ainsi que du sulfure de diméthyle CH3SCH3. Ce dernier est également absorbé par les bactéries marines, mais pas aussi rapidement que le méthanethiol.
Bien que le sulfure de diméthyle constitue généralement moins du quart des produits de dégradation volatils du DMSP, la grande réactivité du méthanethiol abaisse sa concentration à l’équilibre au dixième environ de celle du DMSP (de l’ordre de 0,3 nmol·L-1 contre 3 nmol·L-1 respectivement). Une fraction importante du sulfure de diméthyle marin est oxydée en diméthylsulfoxyde (DMSO) et l’absorption abiotique et microbienne du méthanethiol dans l’eau de mer contribuent à écarter les concentrations relatives de ces deux composés du facteur de 1 à 10 observé à l’équilibre entre le méthanethiol et le sulfure de diméthyle.
À la surface de l’eau de mer, le méthanethiol est le produit principal de la décomposition du métabolite diméthylsulfoniopropionate (DMSP) des algues. Il semblerait que les bactéries marines récupèrent la plupart de leurs protéines soufrées via la décomposition du DMSP et l’incorporation du méthanethiol et non pas par les sulfates. Pourtant ces dernières sont bien plus concentrés dans l’eau de mer (28 nM contre environ 0,3 nM pour le méthanethiol). Les bactéries d’environnements oxiques et anoxiques peuvent également convertir le méthanethiol en diméthylsulfide (DMS), bien que la plupart du DMS en surface de l’eau de mer soit en réalité produit par d’autres mécanismes. Le DMS et le méthanethiol peuvent tous deux être utilisés par certains microbes comme substrats pour la méthanogenèse de certains sédiments anoxiques.
Le sulfure d’hydrogène joue un rôle important en biologie. Il est produit par la dégradation des protéines contenant du soufre et est en grande partie responsable de l’odeur fétide des excréments et des gaz intestinaux, humains et animaux.
Il peut résulter de décomposition bactérienne de la matière organique dans des environnements pauvres en oxygène (méthanisation), ou de l’action des bactéries sulfato-réductrices.
- en mélangeant du soufre et de la limaille de fer, on réalise un brûlât dans un têt (production d’une fumerolle blanche et âcre), ce qui donne une sorte de caillou orangé (le sulfure de fer(II) (FeS) avec des traces grises (la limaille qui n’a pas réagi) ;
- en versant n’importe quel acide (de préférence de l’acide sulfurique, mais c’est l’ion hydronium qu’on fait réagir) sur le sulfure de fer obtenu à la première étape, il se produit immédiatement un dégagement intense de sulfure d’hydrogène bien reconnaissable à son odeur qualifiée dans les manuels scolaires de « nauséabonde », qui est celle de l’œuf pourri.
On peut également faire réagir du sulfure d’aluminium Al2S3 avec de l’eau, ce qui produit, en plus du H2S, de l’hydroxyde d’aluminium.
Dans les couches terrestres, la zone de réduction des sulfates par les microorganismes est comprise entre la zone méthanogène et la zone de réduction du fer lorsqu’une source de carbone existe dans le sol. Le cycle du soufre peut interagir avec ces deux cycles. Le cycle du soufre produit de l’H2S lors de la réduction catabolique du soufre sous forme de SO42−. Le H2S, produit par exemple dans un milieu dulçaquicole, va piéger le fer sous forme de FeS2. Dans ce milieu type, le piégeage du fer va bloquer le cycle du phosphore, car le fer ne pourra plus piéger le phosphate. Le phosphate non-piégé se traduit souvent par une prolifération des algues, et est déterminant dans le phénomène d’eutrophisation.
Le soufre sous sa forme réduite H2S est un donneur d’électron utilisé par les bactéries phototrophes non productrices d’oxygène (ex: les bactéries vertes et bactéries pourpres sulfureuses).
Les bactéries méthanogènes entrent en compétition avec les bactéries réduisant le sulfate : la réduction du SO42− est énergétiquement plus favorable que celle du méthane. Ainsi, en milieu naturel, s’il y a plus de SO42− que de méthane, on observera plutôt un développement des bactéries sulfitoréductrices que des méthanogènes. De plus, les rejets de SO42− par les activités humaines ont une influence sur le cycle du méthane. En effet, SO42− diminue la production de méthane, réduisant ainsi le méthane atmosphérique qui joue un rôle important sur l’augmentation de l’effet de serre. De ce fait, SO42− peut être utilisé pour réduire l’effet de serre.
L’oxydation photolitotrophe est réalisée en milieu anaérobie par les phototrophes (bactéries pourpres sulfureuses et bacteries vertes sulfureuses). Elles utilisent le CO2 comme source de carbone et le H2S comme source d’hydrogène pour la photosynthèse.Ces bactéries font une photosynthèse dite anoxygénique réduisant le CO2 par le H2S. En conditions anaérobies, le H2S cède des électrons au CO2, le produit étant le soufre élémentaire : 2H2S + CO2 → CH2O + H2O +2S.
Cette réaction est endergonique : elle utilise la lumière comme source d’énergie.
La réduction anabolique des sulfates SO42− est utilisée dans la biosynthèse des acides aminés et des protéines. Elle s’effectue en milieu aérobie, soit dans des sédiments ou sols mouillés et oxygénés. Elle est appelée réduction assimilative (ou assimilatrice) par opposition à la réduction catabolique des sulfates, dite dissimilatrice, se déroulant en milieu anaérobie.
La réduction aérobie des sulfates est dite anabolique car elle consomme de l’énergie (sous forme d’ATP ou NADPH + H+) afin de permettre l’incorporation par réduction d’une molécule inorganique (ici le sulfate) dans la matière organique.
Les sulfates sont réduits en sulfures pour permettre la synthèse des acides aminés soufrés (cystéine et méthionine) ou encore de coenzymes. Les plantes, mais également des bactéries sulfatoréductrices aérobies, permettent cette réduction anabolique.
La réduction aérobie des sulfates est très complexe. Elle nécessite son activation par la formation de 3′-phosphoadénosine 5′-phosphosulfate suivie de sa réduction en sulfite SO32−. Puis celui-ci est directement réduit en sulfure d’hydrogène H2S par la sulfite-réductase assimilative7.
La cystéine peut alors être synthétisée à partir de H2S (chez les plantes et les bactéries) en deux étapes, la sérine étant le précurseur de la cystéine :
- Dans un premier temps, l’acétylsérine est formée à partir de l’acétyl-CoA dont le groupement acétyle est transféré à la sérine.
- Dans un second temps, H2S déplace le groupement acétyle pour former la cystéine.
Après sa synthèse, la cystéine participe à l’élaboration d’autres molécules organiques contenant du soufre. Ainsi, le soufre est réintroduit dans la matière organique
https://www.nature.com/articles/s41396-023-01539-1
https://www.nature.com/articles/s41396-021-01047-0
https://www.sciencedirect.com/science/article/abs/pii/S0016703716304860
https://www.sciencedirect.com/science/article/abs/pii/S0009254114000734
https://www.sciencedirect.com/science/article/abs/pii/S0065211320301073
https://www.sciencedirect.com/science/article/abs/pii/S0012825213000512
https://www.sciencedirect.com/science/article/abs/pii/S0883292706000904
https://www.sciencedirect.com/science/article/abs/pii/S0016703711001773
https://www.sciencedirect.com/science/article/abs/pii/S0944501323000423
https://www.sciencedirect.com/science/article/abs/pii/S0269749123015981
https://www.sciencedirect.com/science/article/abs/pii/S0959652623032717
https://www.sciencedirect.com/science/article/abs/pii/S0048969723035210
https://www.sciencedirect.com/science/article/abs/pii/S0045653507007151
https://www.sciencedirect.com/science/article/abs/pii/S1385894723039761
https://www.nature.com/articles/s43705-023-00261-5
https://www.nature.com/articles/ncomms13219
https://www.nature.com/articles/s41396-020-0602-x#Sec8
https://www.nature.com/articles/s41564-019-0527-1
https://www.nature.com/articles/nrmicro2653
https://link.springer.com/article/10.1007/s10533-007-9091-5
https://link.springer.com/article/10.1007/s13280-021-01612-z
https://link.springer.com/article/10.1007/s10533-007-9091-5#ref-CR122
https://royalsocietypublishing.org/doi/10.1098/rspa.2019.0769
https://cdnsciencepub.com/doi/abs/10.1139/f04-052
https://www.nature.com/articles/s41396-019-0573-y
https://www.nature.com/articles/s41467-023-37759-4
https://www.nature.com/articles/s41396-020-00817-6
Protocole de conservation pour l’analyse du diméthylsulfoniopropionate et du diméthylsulfoxyde dans le matériel végétal de l’herbier méditerranéen Posidonia oceanica et réévaluation de la teneur en feuilles de diméthylsulfoniopropionate
https://www.sciencedirect.com/science/article/abs/pii/S0304377017301936 [43]
Le diméthylsulfure (DMS) est ce que principal gaz biogénique sulfuré émis de l’océan vers l’atmosphère. Il peut exercer un effet refroidissant sur le climat formant des aérosols sulfates et intensifiant l’albédo des nuages de basses altitudes, diminue la quantité d’énergie radiative atteindre la surface de la Terre. Bien que le SGD homme être produit directement par certaines espèces phytoplanctoniques, il provient principalement des bactéries hétérotrophes via la dégradation de fils précurseur, le diméthylsulfoniopropionate(DMSP). Le DMSP est synthétisé par le phytoplancton en différentes concentrations selon les conditions environnementales, incluant la disponibilité du fer. La présente étude porte sur la dynamique microbienne du DMSP dans le Pacifique nord-est le long d’un dégradé naturel du fer.
Le diméthylsulfoniopropionate (DMSP) est produit en quantités variables par de nombreux organismes marins dont le phytoplancton, les macroalgues, les herbiers marins, ainsi que des angiospermes terrestres, en particulier certaines plantes des marais salants intertidaux comme certaines espèces de Spartina. Plusieurs rôles physiologiques et écologiques ont été attribués au DMSP, tels qu’osmolyte, osmorégulateur, agent de dissuasion des herbivores, cryoprotecteur, antioxydant contre les espèces réactives de l’oxygène (ROS), métabolite de débordement du carbone, antimicrobien contre les virus ou les bactéries. Le diméthylsulfoxyde (DMSO) est présent dissous dans l’eau de mer, ainsi que de manière intracellulaire dans le phytoplancton et dans les macrophytes intertidaux tels que Spartina. Le DMSO est formé à partir de l’oxydation du sulfure de diméthyle (DMS) dans l’eau de mer par photo-oxydation et dans les cellules, vraisemblablement par piégeage des ROS. Nous avons récemment montré la présence de DMSP chez la phanérogame marine Posidonia oceanica , endémique de la mer Méditerranée, et les variations de la teneur en feuilles de DMSP en fonction de la saison et de la profondeur ont été attribuées à la réponse aux changements de disponibilité de lumière. Nous avons conclu que le rôle physiologique du DMSP chez P. oceanica était d’agir comme antioxydant des ROS liées à une intensité lumineuse élevée, par analogie avec Spartina alterniflora, et comme également montré pour le phytoplancton et les macroalgues.
EFFETS DU STRESS FER ET LUMIERE SUR LA COMPOSITION BIOCHIMIQUE DU PHAEOCYSTIS SP ANTARCTIQUE. (PRYMNÉSIOPHYCÉES). I. CONCENTRATIONS INTRACELLULAIRES DE DMSP
https://onlinelibrary.wiley.com/doi/abs/10.1046/j.1529-8817.1998.340486.x [44]
Le fer est essentiel à la croissance du phytoplancton, car il intervient dans de nombreux processus métaboliques. Il contrôle la photosynthèse ainsi que de nombreux processus enzymatiques. Ainsi, le fer affecte l’approvisionnement énergétique de la cellule et contribue à l’assimilation du carbone et de l’azote. Pour déterminer si une limitation en fer entraînerait un stress énergétique ou une carence en azote induite, une Phaeocystis sp . (Prymnesiophyceae) a été étudiée pour sa composition biochimique, en mettant l’accent sur la production intracellulaire de diméthylsulfoniopropionate (DMSP). Le DMSP est suggéré pour remplacer les solutés contenant de l’azote dans des conditions de carence en azote. Cultures par lots de Phaeocystis sp. ont été cultivées dans des conditions riches et pauvres en fer et simultanément soumises à des intensités lumineuses élevées et faibles. La déplétion en fer a induit une chlorose et a supprimé les taux de croissance ainsi que le rendement maximal des cultures ; ces effets étaient renforcés par de faibles intensités lumineuses. Les volumes cellulaires étaient fortement réduits dans des conditions limitées en fer. Cependant, cette réduction du volume cellulaire s’accompagnait d’une teneur réduite en DMSP uniquement dans les cultures soumises à de faibles intensités lumineuses. Dans des conditions de forte luminosité, aucune réduction du DMSP n’a été observée ; par conséquent, les concentrations intracellulaires de DMSP ont augmenté. Ces observations sont discutées par rapport au métabolisme du carbone et de l’azote et à la voie de biosynthèse du DMSP. Il est avancé que dans des conditions de forte luminosité et de faible teneur en fer, les cellules étaient à la limite d’une carence en azote induite par une limitation en fer, alors que dans des conditions de faible luminosité et de faible teneur en fer, les cellules étaient limitées en énergie, ce qui entraînait une suppression globale des taux métaboliques. Entre les traitements, les ratios DMSP/chlorophylle- a variaient d’un facteur 5, démontrant la dépendance de ce paramètre sur l’état physiologique de la cellule.
Contraintes environnementales sur la production et l’élimination du diméthylsulfure de gaz (DMS) climatiquement actif et implications pour la modélisation des écosystèmes
https://link.springer.com/article/10.1007/s10533-007-9091-5 [45]
Les concentrations dans l’eau de mer du composé sulfuré volatil diméthylsulfure (DMS) qui refroidit le climat sont le résultat de nombreux processus de production et de consommation au sein de l’écosystème marin. En raison de cette nature complexe, il est difficile de prédire les schémas de distribution temporelle et géographique des concentrations de DMS et l’inclusion du DMS dans les modèles climatiques océaniques mondiaux n’a été tentée que récemment.
La description des processus comprend : (1) les paramètres contrôlant la production de DMSP tels que la composition en espèces et les facteurs abiotiques ; (2) la conversion du DMSP en DMS par des enzymes algales et bactériennes ; (3) le sort du DMSP-soufre dû, par exemple, au pâturage, à la consommation microbienne et à la sédimentation et (4) aux facteurs contrôlant l’élimination du DMS de la colonne d’eau, tels que la consommation microbienne, la photo-oxydation et l’émission dans l’atmosphère. Le sulfure de diméthyle (DMS) est un composé organique semi-volatile du soufre qui représente 50 à 60 % du flux total naturel de soufre réduit dans l’atmosphère, y compris les émissions des volcans et de la végétation. En fournissant 95 % du flux vers l’atmosphère, les océans sont la principale source de DMS. À la fin des années 1980, l’hypothèse selon laquelle le DMS serait impliqué dans la régulation biologique du climat global a été avancée. Ce n’est cependant que récemment que le DMS a été incorporé dans les modèles climatiques mondiaux. Après émission dans l’atmosphère, ce composé soufré volatil est oxydé en dioxyde de soufre (SO 2 ) et en d’autres produits. À partir du SO 2 , du sulfate non marin (nss) est produit, qui peut former du sulfate (SO2−4) particules qui agissent comme noyaux de condensation de la vapeur d’eau. Ces noyaux affectent les propriétés radiatives de l’atmosphère et des nuages, avec des implications sur le climat. Un nombre plus élevé de noyaux de condensation détournera davantage de rayonnement solaire entrant vers l’espace et réduira ainsi la température sur Terre. L’hypothèse selon laquelle ce processus pourrait moduler l’effet de serre dû à l’augmentation des apports anthropiques de CO 2 dans l’atmosphère a été indirectement étayée par les résultats de la modélisation de l’effet des apports anthropiques de SO 2 dans l’atmosphère. Bien que cette étude ait donné une indication approximative des effets antagonistes du SO 2 et du CO 2 atmosphériques , une compréhension quantitative de toutes les sources et puits d’aérosols atmosphériques fait encore défaut. Actuellement, la production anthropique de SO 2 dépasse la production naturelle de SO 2 d’un facteur 3, mais l’impact de la première sur la production d’aérosols est largement confiné aux zones industrialisées de l’hémisphère Nord. Par conséquent, l’échange de DMS marins revêt une grande importance régionale et peut affecter le climat à l’échelle mondiale.
Dans l’une des premières tentatives visant à ajouter du DMS dans un modèle climatique océanique mondial, Bopp et al. ont montré qu’un doublement de la concentration atmosphérique en CO 2 entraînait une réduction du flux de DMS aux basses latitudes et un enrichissement aux latitudes moyennes. Ainsi, selon le signe du hasard dans le flux de DMS, le forçage climatique ultérieur par les produits soufrés pourrait soit atténuer, soit amplifier l’effet de serre. L’inclusion du DMS dans le modèle a été réalisée en couplant la production de diméthylsulfoniopropionate (DMSP) et sa conversion en DMS avec l’état trophique de l’écosystème, lui-même basé sur le taux de silice, défini comme la production locale simulée de silice biogénique. par rapport à la production maximale et est liée à la prolifération des diatomées. Elle est calculée à partir des concentrations locales de silice et d’un rapport Si:C variable, qui dépend de la concentration en silice. Bien que simpliste dans son approche écologique, cette étude a montré qu’une augmentation de la concentration de CO 2 n’entraîne pas nécessairement une augmentation de la production de DMS susceptible de contrebalancer l’effet de serre, comme le suggère l’hypothèse de Charlson-Lovelock-Andreae-Warren. De toute évidence, une meilleure compréhension des processus biologiques est nécessaire pour aborder le rôle du DMS dans les mécanismes de rétroaction climatique.
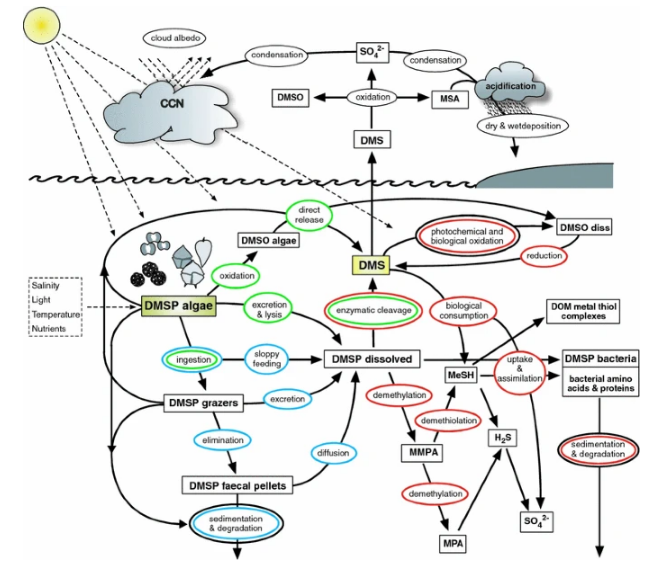
Dans cette revue, une grande attention a été accordée à deux taxons d’haptophytes d’algues spécifiques : Phaeocystis sp. et Emiliania huxleyi . Ces algues sont bien connues comme productrices prolifiques de DMS et de DMSP et leurs proliférations peuvent couvrir de vastes zones dans les eaux néritiques et océaniques. Cependant, il n’existe pas de relation directe puisque la production de DMSP est limitée à un nombre limité de taxons d’algues. Un autre facteur de complication est que les conditions physiologiques des cellules d’algues affectent la production de DMSP. En conséquence, il n’existe pas de relation globale définitive entre les paramètres de la biomasse algale tels que la chlorophylle et le DMSP algal. De nombreux modèles d’écosystèmes sont exprimés en réservoirs de carbone ou d’azote, alors que de nombreux modèles climatiques mondiaux sont plus souvent exprimés en unités de carbone.
Le DMSP est un composé multifonctionnel et il ne fait aucun doute qu’il joue un rôle de soluté compatible dans le métabolisme cellulaire, mais la régulation de sa concentration interne n’est toujours pas résolue.
Étant donné que de nombreux paramètres abiotiques semblent avoir un effet dans une certaine mesure, Stefels a émis l’hypothèse que la production de DMSP pourrait également servir de mécanisme de débordement pour l’excès de soufre réduit dans des conditions de croissance déséquilibrée, lorsque les flux de carbone et d’azote sont désaccordés. La production continue et la perte éventuelle de DMSP serviraient de puits pour l’excès de carbone tout en régénérant l’azote intracellulaire à partir de la méthionine, qui pourrait ensuite être utilisé pour la synthèse d’autres acides aminés. Bien qu’un tel mécanisme semble inutile, les avantages sont le maintien de la machinerie métabolique. Une autre hypothèse sur la fonction physiologique du DMSP a été présentée par Sunda. Il suggère que le DMSP et ses produits de dégradation, le DMS, l’acrylate, le diméthylsulfoxyde (DMSO) et l’acide méthane sulfinique (MSNA), forment ensemble une cascade de capteurs de radicaux qui peuvent servir de système antioxydant efficace qui devrait être réglementé en partie par le clivage enzymatique du DMSP. Il est impossible de mesurer les flux à travers cette cascade de composés. Sunda et ses collègues ont suggéré l’hypothèse des antioxydants sur la base de concentrations élevées de DMSP intracellulaire dans des conditions de stress. Étant donné qu’une croissance déséquilibrée et la production de ROS coexistent souvent dans des conditions d’irradiation élevée et/ou de nutriments limités, il est difficile de tester les deux hypothèses individuellement sans une étude détaillée de l’état physiologique des cellules et des flux à travers les voies biochimiques pertinentes. De plus, les deux hypothèses ne doivent pas nécessairement s’exclure mutuellement, puisqu’une fonction dans la gestion du stress oxydatif n’exclut pas des fonctions supplémentaires dans le métabolisme cellulaire.
Il est clairement montré que la production spécifique de DMS + DMSP est couplée à la croissance cellulaire et se compare bien dans diverses conditions : une gamme de salinités, des conditions d’éclairage faibles ou élevées et une limitation de l’azote ou du phosphore. Un piège potentiel dans les modèles basés sur le carbone réside cependant dans le découplage entre la croissance cellulaire et la croissance du carbone, qui entraînerait des modifications des ratios DMSP:C. Les preuves directes d’un tel découplage entre les cycles du carbone et du soufre font encore défaut, mais des preuves indirectes peuvent être recherchées dans les ratios DMSP:C calculés par rapport aux taux de croissance. Cela peut entraîner une sous-estimation du carbone cellulaire sous limitation de N ou Fe (et donc une surestimation du rapport DMSP:C), puisque, dans ces conditions, les cellules deviennent souvent plus denses en carbone, c’est-à-dire une augmentation du carbone cellulaire par volume cellulaire.
Les algues peuvent ajuster la concentration intracellulaire de DMSP par les voies biosynthétiques (anabolisantes) ou de dégradation (cataboliques). Les enzymes DMSP-lyase facilitent la voie de dégradation, dans laquelle le DMSP est clivé en DMS, en acrylate et en proton. Ce qui contrôle l’activité des DMSP-lyases dans le phytoplancton est encore inconnu.
Le DMSP présent dans les diatomées de glace peut être une source importante de DMSP d et de DMS, suite à la débâcle des glaces dans les eaux polaires, lorsque les communautés planctoniques interstitielles sont libérées des eaux interstitielles hypersalines dans l’eau de mer de salinité beaucoup plus faible. L’autolyse a longtemps été désignée comme la principale voie de libération du DMSP particulaire, puisque, à la fin d’une prolifération, les cellules se désintègrent et le DMSP est libéré dans l’eau. Ce processus est ainsi parallèle à une éventuelle exsudation active par des cellules vivantes limitées en nutriments. Une explication mécaniste de la libération de DMS pendant le pâturage a ensuite été fournie. Wolfe et Steinke ont découvert que le broutage par le dinoflagellé herbivore Oxyrrhis marina , qui engloutit ses proies par phagotrophie, pouvait augmenter considérablement la production de DMS et que le rendement en DMS de la proie ingérée-DMSP était corrélé à l’activité lyase in vitro de la proie.
La demi-vie chimique du DMSP dans l’eau de mer est supérieure à 8 ans, ce qui se traduit par une stabilité abiotique élevée dans des conditions naturelles. Par conséquent, la majeure partie de l’élimination du DMSP se fait par des processus enzymatiques. Dans le réseau alimentaire microbien, le DMSP dissous a de nombreux destins et plusieurs études récentes sur les voies microbiennes et les mécanismes impliqués. En effet, plusieurs études ont montré que le DMSP à lui seul peut contribuer à hauteur de 1 à 15 % à la demande totale de carbone bactérien dans les eaux de surface. De plus, l’assimilation du DMSP peut satisfaire la plupart, sinon la totalité, de la demande en soufre des bactéries marines. Les deux principales voies de dégradation bactérienne du DMSP sont le clivage en DMS et en acrylate et la déméthylation/déméthiolation, avec ou sans absorption préalable dans les cellules. Quantitativement, la voie de dégradation la plus importante du DMSP est la déméthylation. Cette voie ne donne pas de DMS, mais du 3-méthiolpropionate (MMPA) et, après une seconde déméthylation, du 3-mercaptopropionate (MPA). Le MMPA peut également produire du méthanethiol (MeSH) après une réaction de déméthiolation. Outre le DMS, le MeSH est un autre composé soufré volatil important. Il est métabolisé rapidement et semble être la principale source de soufre pour la production d’acides aminés et de protéines soufrés dans le bactérioplancton. Comparé au DMS et au DMSP, le diméthylsulfoxyde non volatil (DMSO) est un composant mal compris du cycle du soufre marin. En plus du fait que le DMSO est omniprésent dans l’eau de mer en phase dissoute, des preuves s’accumulent également pour la production directe de DMSO particulaire par des cultures et des assemblages naturels de microalgues marines.
La dégradation bactérienne du DMS dominait les échanges gazeux mer-air. Les principales voies de dégradation microbienne du DMS sont la consommation via les monooxygénases et les méthyltransférases DMS et l’oxydation via la déshydrogénase DMS. Le DMS peut être oxydé en DMSO par une variété d’oxydants de soufre et d’ammoniac, de méthylotrophes et de phototrophes, ce qui suggère qu’il s’agit d’un processus polyvalent à l’échelle mondiale. Dans les eaux riches en nitrates, 35 % de la photolyse du DMS observée était liée à la photochimie des nitrates et que les taux de photolyse augmentaient de manière linéaire avec l’ajout de nitrate. Cette dépendance aux nitrates a également été observée. Par ailleurs, la photolyse du DMS induite par les nitrates était fortement renforcée par la présence d’ions bromure et, dans une moindre mesure, par les ions bicarbonate/carbonate. Contrairement à ce qui était prévu, la photolyse induite par le bicarbonate/carbonate diminue avec l’augmentation du pH.
Cycle du soufre dans le sol médié par les microbes : impact sur la fertilité des sols, la production agricole et la durabilité environnementale
https://www.sciencedirect.com/science/article/abs/pii/S0944501323000423 [46]
Le soufre contrôle le sort de nombreux éléments géochimiques présents dans les sédiments lacustres , notamment le fer, le phosphore et des oligo-éléments importants pour l’environnement. Nous avons mesuré la spéciation du soufre lié aux eaux interstitielles et aux sédiments (sulfate et sulfures aqueux, soufre élémentaire, monosulfure de fer , pyrite, soufre organique) et les variables géochimiques de soutien (carbone, oxygène, fer) dans les sédiments d’un milieu oxygéné en permanence et d’un bassin saisonnièrement anoxique, en utilisant une combinaison d’analyses d’eau interstitielle, d’extractions séquentielles et d’absorption des rayons X près de la structure de bordure. Un modèle diagénétique précoce à l’état non stable a été développé et calibré sur ce vaste ensemble de données pour aider à comprendre les voies et à quantifier les taux de transformations S. Les résultats suggèrent que la principale source de S dans les sédiments est la sédimentation de l’ester-sulfate organique (RO-SO 3 -H). L’hydrolyse de ces composés fournit une source supplémentaire de sulfate pour l’oxydation microbienne anaérobie de la matière organique sédimentaire, libérant du sulfure dans l’eau interstitielle. Les espèces S réduites liées aux solides s’accumulent sous forme de thiols (R-SH) et de sulfures de fer dans le bassin perpétuellement oxygéné et saisonnièrement anoxique, respectivement. Les enrichissements en sulfure de fer formés au cours des dernières décennies de dépôts atmosphériques élevés de SO 4 sont actuellement en train de se dissoudre. Dans les sédiments du bassin perpétuellement oxygéné, cette réaction entrave l’accumulation d’hydroxyde de Fe (III) (oxy) près de l’interface sédiment-eau.
La biogéochimie du soufre (S) est complexe et d’un grand intérêt en raison de son impact sur de nombreux processus environnementaux fondamentaux, allant de la productivité primaire terrestre et du métabolisme microbien des sédiments, jusqu’à la matière organique (MO). Dans les lacs, le cycle du S dans les sédiments est dynamique spatialement et temporellement, et étroitement couplé à d’autres espèces et processus biogéochimiques, car il participe à un réseau complexe de réactions géochimiques impliquant 8 états de valence, allant de + 6 (par exemple, sulfate, SO 4 2− ), 0 (par exemple, soufre élémentaire, S 0 ), à −2 (par exemple, sulfure, S −II et soufre organique réduit).
La majorité des flux de S vers les lacs proviennent des sols et des zones humides du bassin versant, généralement sous forme de matière organique allochtone contenant du S lié à l’oxygène (par exemple, RO-SO 3 – Groupes H tels que l’ester-SO 4 et d’autres organosulfates) ou S liés au carbone (par exemple, groupes R-SH tels que les acides aminés et autres thiols). Les flux aqueux de S vers les lacs proviennent de la minéralisation de ces matières organiques , de l’altération des roches ou du dépôt atmosphérique direct de SO 4 2− . En entrant dans la colonne d’eau, le S organique (S org ) peut être utilisé par les micro-organismes via des enzymes qui clivent les liaisons RO-SO 3 -H ( l’aryl sulfatase) et R-SH ( la cystéine liase), reconstituant ainsi SO 4 2− et HS − , respectivement. Les sources de S à la surface des sédiments comprennent le S dérivé du bassin versant, le SO 4 2− de la colonne d’eau ainsi que l’organisation S se déposant avec le seston. Dans les sédiments, une fois le dioxygène épuisé, le SO 4 2− est principalement réduit en sulfure par réduction dissimilatrice, à partir de laquelle les bactéries sulfato-réductrices gagnent de l’énergie pour leur croissance et leur entretien.
Le sulfure réagit avec Fe pour former du monosulfure amorphe en pyrite cristalline (FeS 2(s) ). La transformation de FeS (m)s en FeS 2(s) est influencée par divers facteurs, notamment le rapport entre Fe réactif et S −II disponible, l’inhibition par carbone organique labile C org , la capacité du S 0 (aq) à favoriser la pyritisation via la voie des polysulfures et l’effet du pH sur la cinétique de transformation du FeS m(s). Globalement, les interactions entre S et Fe dans les eaux anoxiques, les sols gorgés d’eau et les sédiments ont été largement étudiées. Le sulfure peut également être séquestré sous forme de S org, par exemple sous forme de R-SH lors de la sulfuration de la OM et de la réduction assimilatoire du SO 4 2− . L’incorporation de sulfures dans la MO a été fréquemment rapportée dans les sédiments marins, mais moins dans les lacs malgré des preuves de concentrations élevées de thiols dans les sédiments d’eau douce. Les sulfures peuvent en outre être oxydés pour former des espèces S 0 , abondantes à la fois dans les phases solides et aqueuses. S 0 réagit avec Fe et les métaux, et sert de précurseur à la formation de composés S org , y compris des espèces volatiles.
En reconnaissant l’importance cruciale du S dans le contrôle du devenir des nutriments et des éléments majeurs et traces, notre objectif est d’améliorer notre compréhension quantitative de la spéciation et du devenir du S dans les sédiments d’eau douce. En effet, une intégration plus complète du cycle S dans les modèles diagénétiques constitue un tremplin pour une prédiction plus précise des cycles biogéochimiques d’autres éléments, tels que C, O, Fe et P. Nous combinons un vaste ensemble de données sur la spéciation du S en phase aqueuse et solide, recueillies, avec un modèle de transport réactif mis à jour pour S, C, O et Fe. diagenèse. Nous estimons les taux de transformation du S organique et inorganique dans les sédiments de deux bassins lacustres oligotrophes connaissant des conditions d’oxygénation différentes tout au long de l’année, nous permettant de quantifier les voies de séquestration du S dans des conditions oxiques et anoxiques.
Cycle microbien des composés organiques volatils du soufre
https://pubmed.ncbi.nlm.nih.gov/12022467/ [47]
Le cycle microbien des composés organiques volatils du soufre (VOSC), en particulier le sulfure de diméthyle (DMS) et le méthanethiol (MT), est étudié de manière intensive car ces composés jouent un rôle important dans les processus de réchauffement climatique, de précipitations acides et du cycle mondial du soufre. Les concentrations de VOSC dans les sédiments d’eau douce sont faibles en raison de l’équilibre entre la formation et la dégradation de ces composés. Ces réactions se produisent pour la plupart à l’interphase oxique/anoxique des sédiments et de la colonne d’eau. Contrairement aux écosystèmes marins, où le diméthylsulfoniopropionate est le principal précurseur du MT et du DMS, dans les écosystèmes d’eau douce, les VOSC sont formés principalement par la méthylation du sulfure et, dans une moindre mesure, par la dégradation des acides aminés contenant du S. L’une des principales voies de formation de DMS et de MT par méthylation des sulfures est la O-déméthylation anaérobie des composés aromatiques méthoxylés. Des études d’inhibition ont révélé que la majeure partie du MT et du DMS produits de manière endogène est dégradée de manière anaérobie par les méthanogènes.
Facteurs microbiens de la réduction du DMSO et de la méthanogenèse dépendante du DMS dans les sédiments des marais salants
https://www.nature.com/articles/s41396-023-01539-1
Les marais salants sont des environnements très productifs, présentant de grandes abondances de composés organosulfurés. Le diméthylsulfoniopropionate (DMSP) est produit en grande quantité par les algues, les plantes et les bactéries et est un précurseur potentiel du diméthylsulfoxyde (DMSO) et du diméthylsulfure (DMS). Le DMSO sert d’accepteur d’électrons pour la respiration anaérobie conduisant à la formation de DMS, qui est soit émis, soit dégradé par les procaryotes méthylotrophes. Les principaux produits de ces réactions sont des gaz traces à forçage radiatif positif (CO 2 , CH 4 ) ou négatif (DMS) ayant des effets contrastés sur le climat mondial. Ici, nous avons étudié le cycle du soufre organique dans les sédiments des marais salants et suivi la réduction du DMSO dans des expériences par lots anoxiques. Par rapport aux mesures précédentes dans les eaux marines, les concentrations de DMSO dans les sédiments des marais salants étaient jusqu’à environ 300 fois plus élevées. Dans des expériences par lots, le DMSO a été réduit en DMS et ensuite consommé avec une production concomitante de CH 4 . Les changements dans les communautés procaryotes et le nombre de gènes de DMSO réductase ont indiqué une dominance d’organismes contenant les DMSO réductases de type Dms (par exemple, Desulfobulbales , Enterobacterales ). En revanche, lorsque la réduction des sulfates était inhibée par le molybdate, les DMSO réductases de type Tor (par exemple, les Rhodobacterales ) augmentaient. Les vibrionales ont augmenté en abondance relative dans les deux traitements. Des rapports de conversion molaire <1,3 CH 4 par DMSO ajouté étaient accompagnés d’une prédominance des méthanogènes méthylotrophes Methanosarcinales . L’enrichissement des gènes mtsDH , codant pour les méthyl transférases DMS dans les métagénomes, indique leur rôle dans la méthanogenèse dépendante du DMS. Les MAG affiliés à Méthanolobus portaient l’ensemble complet des gènes codant pour les enzymes de la méthanogenèse méthylotrophique.
Les marais salants sont des écosystèmes hautement productifs importants pour la protection côtière, le cycle biogéochimique et la nutrition humaine. Ils sont souvent caractérisés par des zones distinctes en fonction des fréquences d’inondation et des communautés végétales, affectant la composition de leurs nutriments et de leurs communautés microbiennes. La majeure partie de la biomasse produite dans les marais salants est reminéralisée dans les sédiments. Alors que la majorité de la dégradation de la matière organique dans les sédiments marins (~ 50 %) est due à une réduction dissimilatrice des sulfates, d’autres accepteurs d’électrons énergétiquement plus favorables, tels que le DMSO, peuvent également jouer un rôle clé. La réduction respiratoire du DMSO dans des conditions anoxiques produit du diméthylsulfure (DMS), un gaz trace qui agit comme un précurseur des aérosols organiques secondaires, qui peuvent avoir un effet de refroidissement du climat. Un précurseur du DMS et du DMSO est le DMSP, produit par les plantes, les algues et les bactéries. Ainsi, la forte prévalence de ces organismes producteurs de DMSP dans les marais salants favorise le cycle du soufre organique. Des travaux antérieurs ont démontré que l’ajout de DMSO aux boues de sédiments anaérobies des marais salants conduisait à la production de DMS, qui à son tour était consommé avec la production concomitante de CH 4. Le cyclage du DMSO dans les sédiments des marais salants affecte ainsi la production de composés à forçage radiatif positif (CO 2 et CH 4 ) et négatif (DMS). Pour leur petite superficie mondiale d’environ 0,01 %, les marais salants contribuent de manière surproportionnée (~ 0,28 %) aux émissions mondiales de DMS et de CH 4 marin (~ 13 %). Comprendre le sort du DMS et du DMSO dans ces écosystèmes séquestrant le carbone (carbone bleu) est donc important pour modéliser leur effet sur le climat, ainsi que le renouvellement du carbone et du soufre.
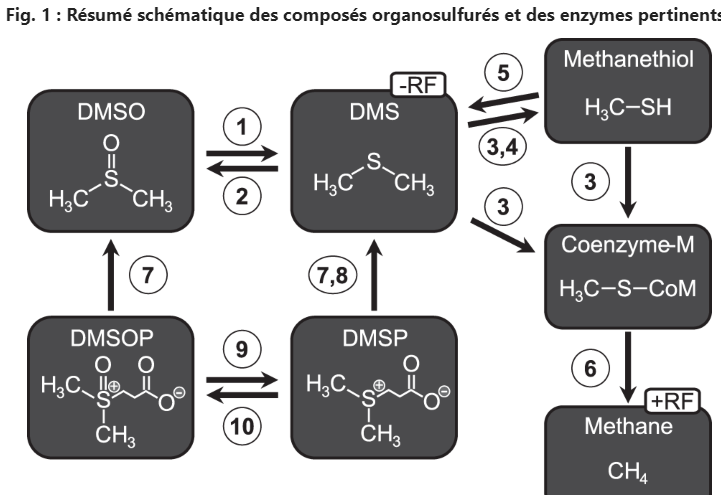
La réduction du DMSO en DMS dans les expériences d’incubation a été suivie d’une diminution des concentrations de DMS et d’une formation simultanée de CH 4. Par conséquent, la majeure partie du DMS provenant de la réduction du DMSO a été utilisée pour la méthanogenèse dépendante du DMS. On s’attendait à ce que la dégradation du DMS en CH 4 génère des composés soufrés inorganiques réduits, tels que le sulfure d’hydrogène (H 2 S), l’abondance relative des Thiotrichales et Thiomicrospirales oxydant le soufre a augmenté. Cela suggère que la concentration de DMSO et la production de DMS qui en résulte ont affecté la compétition pour le DMS entre les méthanogènes et les autres dégradeurs de DMS. La méthanogenèse dépendante du DMS s’est accompagnée d’une augmentation significative de l’abondance relative des Methanosarcinales dans les ensembles de données génétiques de l’ARNr 16S.
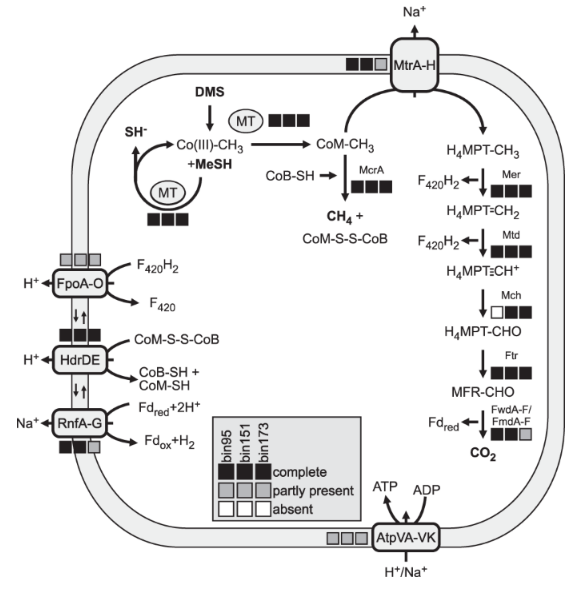
En raison de la pertinence des composés organiques soufrés pour le cycle du carbone et du soufre, les processus individuels, les organismes et les gènes impliqués dans la conversion aérobie du DMS ont été étudiés depuis des décennies. La plupart des études moléculaires indépendantes de la culture se sont concentrées sur le cycle du DMS et du DMSP dans le domaine marin, et seules quelques études ont également envisagé la réduction microbienne du DMSO. La réduction respiratoire du DMSO est connue depuis longtemps comme une source de DMS, qui à son tour peut être émis dans l’atmosphère ou dégradé microbien en CH 4 ou CO 2. Les effets opposés du gaz trace DMS, qui refroidit le climat, et du gaz à effet de serre CH 4 nécessitent une compréhension globale de leur devenir dans l’environnement. La succession observée de métabolites dans nos expériences d’incubation correspondait aux modèles trouvés dans des études précédentes. Sur la base des concentrations de DMS et de CH 4 dans l’espace libre des incubations de suspension, le processus peut être séparé en deux phases distinctes : la réduction initiale du DMSO avec production de DMS suivie par la dégradation du DMS avec méthanogenèse. Bien que ces phases aient été stimulées par l’ajout de DMSO pour enrichir les organismes à l’origine de ces processus, elles se chevauchent dans les incubations et se produisent très probablement simultanément dans la nature ou potentiellement dans les régions voisines du profil sédimentaire en milieu naturel. Le DMSP est considéré comme la principale source de DMS dans les environnements marins. Nos observations indiquent que le DMSP et le DMSO sont facilement disponibles dans les sédiments des marais salants, où une communauté diversifiée de bactéries hétérotrophes les utilise.
Les résultats de nos expériences d’incubation suggèrent un effet négatif du molybdate sur l’abondance relative du SRB et de la plupart des autres bactéries porteuses du dmsA . Seuls quelques groupes de bactéries porteuses de dmsA (par exemple, Zoogloeaceae , Sporomusaceae ) ont augmenté en abondance relative dans DMSO + Mo mais ont diminué dans les traitements au DMSO. Les concentrations élevées de DMSO dans les marais salants mettent en évidence son importance pour la respiration anaérobie et la production ultérieure de DMS. Nous avons découvert un potentiel génétique répandu pour la réduction du DMSO dans l’environnement, avec une forte prévalence du type Dms du DMSOR. L’augmentation relative des méthanosarcinacées porteuses de mtsDH sous des concentrations élevées de DMS montre le rôle du DMS en tant que substrat non compétitif pour les méthanogènes méthylotrophes. Cependant, la méthanogenèse en tant que puits de DMS pourrait être plus faible dans des conditions naturelles et devra être quantifiée dans les recherches futures. L’effet supposé de refroidissement climatique des émissions de DMS serait annulé par la transformation microbienne en gaz à effet de serre CH 4 et CO 2 .
Le tensioactif améliore la production de sulfure d’hydrogène par fermentation anaérobie : modifications de la structure des matières organiques contenant du soufre et de la communauté microbienne
https://www.sciencedirect.com/science/article/abs/pii/S0048969723016431
La présence de tensioactifs dans le système de boues activées (WAS) est généralement considérée comme bénéfique pour le traitement des boues , notamment en améliorant la déshydratation des boues et la génération de produits de fermentation à valeur ajoutée. Cependant, dans cette étude, il a été découvert pour la première fois que le dodécylbenzène sulfonate de sodium (SDBS, un tensioactif typique) augmentait manifestement la production de sulfure d’hydrogène (H 2 S) toxique à partir de la fermentation anaérobie WAS à des concentrations pertinentes pour l’environnement. Les résultats expérimentaux ont montré que la production de H 2 S à partir du WAS augmentait de manière significative, passant de 53,24 × 10 −3 à 111,25 × 10 −3 mg/g de matières volatiles en suspension (VSS) lorsque le niveau de SDBS augmentait de 0 à 30 mg/g de solides en suspension totaux (TSS). Il a été constaté que la présence de SDBS détruisait la structure du WAS et augmentait la libération de matières organiques contenant du soufre. Le SDBS a réduit la proportion de structure en hélice α, endommagé les ponts disulfure et la conformation des protéines et détruit efficacement la structure des protéines. Le SDBS a favorisé la dégradation des matières organiques contenant du soufre et a fourni des micromolécules organiques plus facilement hydrolysées pour la production de sulfures. L’analyse microbienne a montré que l’ajout de SDBS augmentait l’abondance de gènes fonctionnels codant pour la protéase, les transporteurs de cassettes de liaison à l’ATP et la lyase d’acides aminés, augmentait les activités et l’abondance des microbes hydrolytiques, augmentant ainsi la production de sulfures provenant de l’hydrolyse de matières organiques contenant du soufre. Par rapport au témoin, 30 mg/g de TSS SDBS ont augmenté l’hydrolyse des soufres organiques et la dégradation des acides aminés de 47,1 % et 63,5 %, respectivement. L’analyse des gènes clés a en outre montré que l’ajout de SDBS favorisait le système de transport des sulfates et la réduction dissimilatoire des sulfates. La présence de SDBS a également abaissé le pH de fermentation, favorisé la transformation à l’équilibre chimique du sulfure, augmentant ainsi la libération de gaz H 2 S.
La fermentation anaérobie est considérée comme une méthode prometteuse pour traiter les boues activées résiduaires (WAS), au cours de laquelle les boues sont réduites, les agents pathogènes sont tués et l’énergie et les ressources de grande valeur (par exemple, les acides gras à chaîne courte (SCFA) et l’hydrogène) sont récupérées. Cependant, le faible rendement et la production de gaz H 2 S toxique et malodorant limitent l’application à grande échelle de la technologie de fermentation anaérobie des boues. Par exemple, l’efficacité globale de dégradation des solides volatils (VS) dans les WAS est généralement inférieure à 30 % au cours du processus de fermentation anaérobie. Pendant ce temps, des niveaux mortels de H 2 S (c’est-à-dire ∼ 5 000 à 8 000 ppm) sont fréquemment détectés dans les cuves de fermentation WAS, tandis que même de faibles niveaux de H 2 S (par exemple 20 à 100 ppm) nuisent à la santé humaine, corrodent l’équipement de fermentation et endommage les micro-organismes de fermentation.
Au cours des dernières années, de nombreux efforts ont été consacrés à l’amélioration de l’efficacité de la fermentation WAS et se sont révélés efficaces pour favoriser la désintégration des boues et augmenter la génération de produits de valeur. En comparaison, moins d’efforts ont été consacrés à l’étude de la génération de H 2 S provenant de la fermentation des boues. La plupart des publications antérieures dans ce domaine se sont concentrées sur le développement de méthodes de prétraitement pour éliminer la production de H 2 S. Par exemple, il a été rapporté que le prétraitement électrochimique à 12 V pendant 720 s réduisait 99 % du sulfure dissous et 100 % du gaz H 2 S pendant le traitement des boues. Bien que toutes ces méthodes soient efficaces pour éliminer le H 2 S de la fermentation anaérobie dans une certaine mesure, les paramètres opérationnels optimaux et les efficacités d’élimination diffèrent évidemment, ce qui dépend généralement des propriétés et de la composition des boues. À ce jour, aucun effort n’a été fait pour explorer l’impact de la composition des boues sur la génération de H 2 S issue de la fermentation anaérobie WAS.
En plus de certaines matières organiques disponibles (par exemple, protéines et glucides), les WAS, en tant que sous-produit majeur des STEP, contiennent également de nombreuses autres substances telles que des contaminants émergents, des nanoparticules et des tensioactifs. Le dodécylbenzène sulfonate de sodium, en tant que tensioactif largement utilisé, a été appliqué dans les produits ménagers et industriels. Après l’utilisation d’un produit à base de SDBS (par exemple un détergent), le SDBS est généralement rejeté dans les égouts, collecté dans les STEP, absorbé et accumulé dans les boues activées en raison de sa forte nature hydrophobe. Jiang et coll. ont constaté que le niveau de SDBS dans les boues atteignait 31 mg/g de poids sec. Étant donné que WAS contient du SDBS à haute concentration, plusieurs publications ont été exploitées pour étudier son effet sur le traitement WAS. Presque toutes les publications précédentes ont montré que l’existence du SDBS était bénéfique pour la fermentation anaérobie des boues et augmentait la génération de produits de fermentation à valeur ajoutée tels que les SCFA et le H 2 . Par exemple, il a été rapporté qu’avec une augmentation de la teneur en SDBS de 0 à 20 mg/g de TSS, le plus grand rendement en SCFA augmentait de 339,1 à 2 599,1 mg DCO/L (Jiang et al., 2007a). De plus, de nombreuses méthodes de traitement basées sur l’ajout de tensioactifs ont été proposées et appliquées à la fermentation WAS. Ces résultats ont indiqué que l’existence de tensioactifs dans le WAS ou l’ajout direct de tensioactifs au WAS est utile à la fermentation anaérobie du WAS.
Malgré quelques progrès réalisés dans la littérature précédente, l’influence du SDBS sur la production de gaz H 2 S toxique et malodorant à partir de la fermentation anaérobie des boues par hydrolyse du soufre organique et réduction des sulfates inorganiques, ainsi que sur la migration et la transformation des éléments soufrés issus de la fermentation des boues, restent floues. Le but de cet article était de dévoiler comment le SDBS affecte la génération de H 2 S issue de la fermentation des boues. Tout d’abord, l’impact du SDBS à différents niveaux allant de 0 à 40 mg/g TSS sur la génération de H 2 S provenant de la fermentation des boues a été exploré. Ensuite, les mécanismes par lesquels le SDBS affecte la génération de H 2 S ont été révélés en évaluant les changements dans les voies d’hydrolyse du soufre organique et de réduction du sulfate inorganique. Enfin, l’impact du SDBS sur la communauté microbienne dans la fermentation des boues a également été analysé grâce au fonctionnement de fermenteurs semi-continus. Les résultats obtenus dans cette étude approfondissent la compréhension du rôle des tensioactifs dans la fermentation anaérobie WAS.
L’existence de SDBS a considérablement augmenté la production de SCFA précieux et de H 2 provenant de la fermentation des boues, selon des rapports précédents, ce qui a été davantage étayé dans cet article. Lorsque la teneur en SDBS est passée de 0 à 30 mg/g-TSS, la production maximale d’AGCC a augmenté de 450,3 à 2 584,9 mg DCO/L, tandis que la production cumulée d’hydrogène a augmenté de 2,68 à 11,8 mL/g VSS (Fig. S1). Cependant, il a été constaté que la présence de SDBS augmentait évidemment le taux de H 2 S. La plus grande quantité de petites molécules soufrées facilement hydrolysées contenant des matières organiques et les activités plus élevées des producteurs de H 2 S étaient deux raisons majeures qui favorisaient la production de H 2 S à partir de la fermentation anaérobie du WAS contenu dans le SDBS. Selon la littérature, le tensioactif soluble dans l’eau formerait un complexe organique-surfactant avec les matières organiques (par exemple, phospholipide et protéine) à la surface du WAS, à partir duquel les matières organiques entreraient dans la phase aqueuse sous la forme du complexe. Dans cette étude, l’effet du tensioactif sur la génération de H 2 S fermentaire anaérobie a d’abord été étudié et les mécanismes sous-jacents ont été dévoilés. Les principales conclusions sont :
(1)La présence de SDBS a largement amélioré la génération de H 2 S provenant de la fermentation des boues. Il a été observé que la concentration environnementale de SDBS (c’est-à-dire 30 mg/g de TSS) augmentait la production de H 2 S de 53,204 × 10 −3 à 111,25 × 10 −3 mg/g de VSS.
(2)La présence de SDBS a induit l’extension de la chaîne moléculaire de la protéine, endommageant le disulfure.
Biochimie, physiologie et biotechnologie des bactéries sulfato-réductrices
https://www.sciencedirect.com/science/article/abs/pii/S0065216409012027
Les activités importantes pour l’environnement affichées par SRB sont une conséquence des composants uniques de transport d’électrons ou de la production de niveaux élevés de H 2 S. La capacité de SRB à utiliser des hydrocarbures dans des cultures pures et des consortiums a abouti à l’utilisation de ces bactéries pour la bioremédiation des BTEX (benzène , toluène, éthylbenzène et xylène) dans les sols contaminés. Des souches spécifiques de SRB sont capables de réduire le 3-chlorobenzoate, les chloroéthènes ou les composés nitroaromatiques, ce qui a donné lieu à des propositions visant à utiliser le SRB pour la biorestauration d’environnements contenant du trinitrotoluène et des polychloroéthènes. Étant donné que les SRB ont affiché une réduction dissimilaire de l’U (VI) et du Cr (VI), plusieurs procédures biotechnologiques ont été proposées pour utiliser les SRB dans la bioremédiation des métaux toxiques. Une activité supplémentaire non spécifique sur les métaux réductases a abouti à l’utilisation du SRB pour la récupération des métaux précieux (par exemple le platine, le palladium et l’or) à partir des flux de déchets. Étant donné que le sulfure produit par des bactéries contribue à l’acidification des champs de pétrole, à la corrosion du béton et à la décoloration de la pierre, il existe un intérêt considérable pour le contrôle de l’activité sulfidogénique du SRB. La production de biosulfure par SRB a conduit à l’immobilisation de métaux toxiques et à la réduction des colorants textiles, bien que le processus reste non résolu, les SRB jouent un rôle dans l’oxydation anaérobie du méthane qui non seulement contribue aux activités du cycle du carbone mais épuise également une importante réserve d’énergie industrielle.
L’étude des bactéries sulfato-réductrices (SRB) est passée de la recherche d’une solution à un problème pratique. En recherchant un procédé pour éliminer le sulfate de calcium de l’eau des canaux hollandais et réduire le niveau de minéraux de l’eau destinée à être utilisée dans les chaudières à vapeur, MW Beijerinck a découvert l’activité biologique du « ferment sulfuré » que nous appelons maintenant « réduction dissimilatoire du sulfate ». Beijerinck (1895) a publié plusieurs articles décrivant les caractéristiques et l’activité de son Spirillum desulfuricans nouvellement isolé. Au milieu du XXe siècle, les effets économiques du SRB étaient reconnus dans les domaines suivants : pollution et toxicité attribuées à la bioproduction de H 2 S, biocorrosion des métaux, détérioration des aliments due aux thermophiles producteurs d’endospores et dépenses financières dans l’industrie pétrolière. attribuée aux réducteurs de sulfate. Le rôle du SRB en biotechnologie a été discuté dans des études précédentes. Il est intéressant de noter que, même aujourd’hui, les SRB revêtent une importance économique dans le développement de la biocorrosion et du biofilm dans les champs pétroliers. Au cours des dernières décennies, le développement de nouvelles technologies de culture d’anaérobies et de caractérisation moléculaire des souches a permis de caractériser de nombreux nouveaux genres et espèces.
Dans la réduction dissimilatoire des sulfates (également appelée « respiration des sulfates »), les micro-organismes utilisent le sulfate inorganique comme accepteur d’électrons externe dans l’oxydation des substrats énergétiques, ce qui entraîne la production de sulfure d’hydrogène. Le sulfate peut être utilisé comme accepteur d’électrons terminal à la fois par les membres des bactéries et par les archées. La plupart des procaryotes dotés d’une capacité sulfato-réductrice sont des bactéries, ce qui conforte notre décision d’utiliser le terme SRB au lieu de procaryotes sulfato-réducteurs.
Un cadre biochimique pour l’oxydation anaérobie du méthane pilotée par la respiration dépendante du Fe (III)
https://www.nature.com/articles/s41467-018-04097-9 [48]
La consommation de méthane par les microbes aérobies et anaérobies régit le niveau atmosphérique de ce puissant gaz à effet de serre. Alors que la compréhension biochimique de la méthanotrophie aérobie est bien développée, une compréhension mécaniste de la méthanotrophie anaérobie a été empêchée par l’indisponibilité de cultures pures. Nous rapportons ici une étude biochimique de Methanosarcina acetivorans , une espèce productrice de méthane capable de croissance méthanotrophique anaérobie dépendante de la réduction de Fe (III). Nos résultats soutiennent une voie ancrée par des mécanismes dépendants du Fe (III) pour la conservation de l’énergie conduisant à des réactions endergoniques qui sont essentielles à la croissance méthanotrophique. La production et la consommation de méthane sont une composante du cycle mondial du carbone. Produit final de la décomposition anaérobie de la biomasse, près d’un milliard de tonnes de méthane sont produites chaque année dans divers habitats anaérobies de la biosphère terrestre de la Terre. Le méthane est également produit dans les sédiments marins anaérobies et libéré par de vastes réservoirs d’hydrates de gaz. Le méthane est un gaz à effet de serre important, près de 20 fois plus puissant que le CO 2. Heureusement, les rejets dans l’atmosphère sont atténués par l’oxydation aérobie et anaérobie du méthane ou par son assimilation dans la biomasse cellulaire par les microbes qui contrôlent en fin de compte le climat de la Terre.
L’oxydation anaérobie du méthane (AOM) nécessite une réduction des accepteurs d’électrons (Fe(III), Mn(IV), nitrate ou sulfate) pour être thermodynamiquement favorable. Jusqu’à récemment, on pensait que l’AOM dans les sédiments marins nécessitait une symbiose d’archées anaérobies méthanotrophes (ANME) et d’espèces sulfato-réductrices pour lesquelles ces dernières utilisent le réducteur produit par les premières pour rendre la réaction globale thermodynamiquement favorable. La voie biochimique proposée élève l’AOM de M. acetivorans à un niveau mécaniste fondamental de compréhension. La voie est essentiellement une inversion des voies méthanogènes acétoclastiques et de réduction du CO 2 caractérisées biochimiquement , bien que dépendantes de la réduction du Fe(III). Les résultats révèlent le rôle que joue le Fe (III) dans la conservation de l’énergie et dans la conduite des réactions endergoniques qui soutiennent la croissance méthanotrophique.
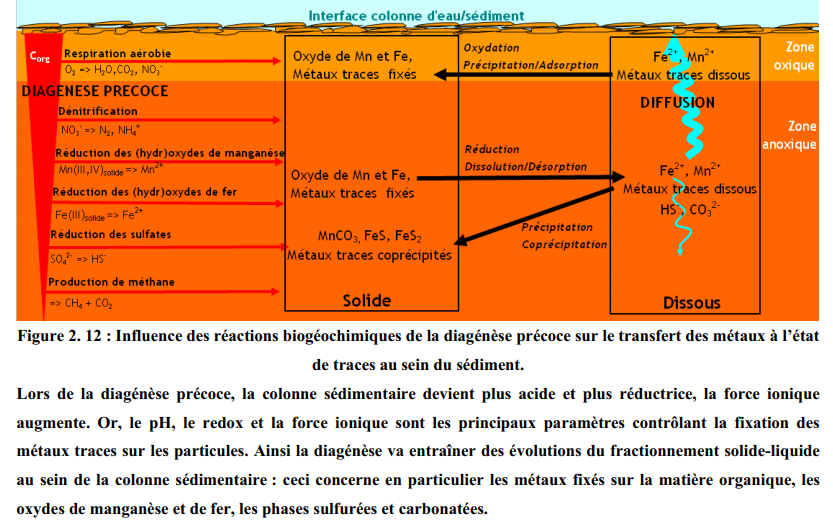
Oxydation simultanée des sulfures et du méthane par un extrémophile
https://www.nature.com/articles/s41467-023-38699-9 [49]
Le sulfure d’hydrogène (H 2 S) et le méthane (CH 4 ) sont produits dans des environnements anoxiques par réduction des sulfates et décomposition de la matière organique. Les deux gaz diffusent vers le haut dans les zones oxiques où les méthanotrophes aérobies atténuent les émissions de CH 4 en oxydant ce puissant gaz à effet de serre. Bien que les méthanotrophes, dans une myriade d’environnements, rencontrent du H 2 S toxique, on ignore pratiquement comment ils sont affectés. Ici, grâce à une culture chimiostatique approfondie, nous montrons qu’un seul micro-organisme peut oxyder CH 4 et H 2 S simultanément à des taux également élevés. En oxydant le H 2 S en soufre élémentaire, le méthanotrophe thermoacidophile Méthylacidiphilum fumariolicum SolV atténue les effets inhibiteurs du H 2 S sur la méthanotrophie. La souche SolV s’adapte à l’augmentation du H 2 S en exprimant une oxydase terminale de type ba 3 insensible aux sulfures et se développe en tant que chimiolithoautotrophe en utilisant le H 2 S comme seule source d’énergie. Des études génomiques ont révélé des enzymes putatives oxydant les sulfures chez de nombreux méthanotrophes, ce qui suggère que l’oxydation du H 2 S est beaucoup plus répandue chez les méthanotrophes qu’on ne le pensait auparavant, leur permettant de relier les cycles du carbone et du soufre de manière nouvelle.
Le sulfure d’hydrogène (H 2 S) est la forme la plus réduite de soufre (S) et une puissante source d’énergie et de soufre, un toxique et une molécule de signalisation. C’est un acide faible qui diffuse facilement à travers les membranes et inhibe divers processus tels que la respiration aérobie en se liant aux cytochromes c oxydases. De plus, d’autres processus métaboliques qui utilisent des enzymes contenant du cuivre et du fer sont sévèrement inhibés par H 2 S. Par conséquent, les micro-organismes vivant dans des environnements riches en sulfures nécessitent des mécanismes adéquats pour détoxifier le H 2 S. Dans une myriade d’environnements, tels que les zones humides, les sédiments marins, le sol, les usines de traitement des eaux usées, les lacs, les rizières, les décharges et les environnements géothermiques acides, le H 2 S est produit par réduction des sulfates (SO 4 2− ), minéralisation de la matière organique. Lors de l’épuisement du sulfate, la matière organique est finalement convertie en méthane (CH 4 ) dans les écosystèmes appauvris en oxygène. Lorsque H 2 S et CH 4 diffusent dans les zones oxiques superposées, le CH 4 peut être utilisé comme source d’énergie par des bactéries aérobies oxydant le méthane, censées atténuer la plupart des émissions de ce puissant gaz à effet de serre.
Il devient de plus en plus clair que les méthanotrophes sont métaboliquement polyvalents et capables d’utiliser des sources d’énergie pertinentes pour l’environnement telles que le H 2 , le propane, l’éthane, l’acétate, l’acétone, le 2-propanol et l’acétol. La possibilité d’utiliser diverses sources d’énergie est très bénéfique dans les environnements où les émissions de gaz sont fortement fluctuantes. Récemment, il a été démontré que des cultures pures du méthanotrophe verrucomicrobien Méthylacidiphilum fumariolicum SolV peuvent consommer du méthanethiol (CH 3 SH), avec la formation sous-stoechiométrique concomitante de H 2 S, indiquant que la souche SolV métabolise partiellement le H 2 S toxique . Par la suite, une étude élégante a démontré que les méthanotrophes protéobactériennes peuvent également oxyder le H 2 S. Les auteurs ont isolé la souche HY1 d’alphaproteobacterium polyvalente « Méthylovirgula thiovorans » d’une tourbière sud-coréenne qui pouvait se développer sur du thiosulfate (S 2 O 3 2− ), du tétrathionate (S 4 O 6 2− ), du soufre élémentaire (S 0 ) et un gamme de composés carbonés. Cependant, les cellules de la souche HY1 cultivées sur CH 4 comme seule source d’énergie n’étaient pas capables d’oxyder le H 2 S, et l’oxydation du H 2 S n’a été initiée et observée que dans les cellules cultivées en présence de thiosulfate. De plus, la croissance sur H 2 S n’a pas été étudiée. Compte tenu des observations récentes, il est primordial d’étudier s’il existe des microbes capables d’oxyder simultanément les gaz importants pour l’environnement CH 4 et H 2 S, comment les méthanotrophes réagissent au H 2 S et si ces méthanotrophes peuvent conserver l’énergie et produire de la biomasse en utilisant le H 2 S comme agent. source d’énergie.
Ici, grâce à une culture chimiostatique étendue, nous montrons pour la première fois qu’un micro-organisme peut oxyder simultanément le CH 4 et le H 2 S. M. fumariolicum SolV est inhibé par la présence de concentrations élevées de H 2 S, mais le H 2 S est rapidement oxydé en soufre élémentaire (S 0 ) comme mécanisme de détoxification pour atténuer l’effet inhibiteur du H 2 S sur l’oxydation du CH 4 . La souche SolV s’adapte au H 2 S avec la régulation positive d’une sulfure de type III : quinone oxydoréductase (SQR) et d’une cytochrome c oxydase de type ba 3 insensible au H 2 S , créant une voie de transfert d’électrons de H 2 S à O 2 . De plus, la souche SolV incorpore CO 2 en utilisant H 2 S comme seule source d’énergie. Nous proposons que la capacité d’oxydation du H 2 S des méthanotrophes verrucomicrobiens est essentielle pour prospérer dans les écosystèmes géothermiques acides riches en soufre. De plus, nous avons trouvé du SQR dans une pléthore de méthanotrophes protéobactériennes de divers environnements. Étant donné que CH 4 et H 2 S coexistent dans une myriade d’écosystèmes limités en oxygène, l’oxydation du H 2 S pourrait être un trait présent chez de nombreux méthanotrophes aérobies.
Méthylation aérobie du sulfure d’hydrogène en sulfure de diméthyle dans divers micro-organismes et environnements
https://www.nature.com/articles/s41396-023-01430-z [50]
Le sulfure de diméthyle (DMS) est la principale source de biosoufre émis dans l’atmosphère et joue un rôle clé dans le cycle mondial du soufre et potentiellement dans la régulation du climat. On pense que le principal précurseur du DMS est le diméthylsulfoniopropionate. Cependant, le sulfure d’hydrogène (H 2 S), un volatile largement répandu et abondant dans les environnements naturels, peut être méthylé en DMS. Les micro-organismes et les enzymes qui convertissent le H 2 S en DMS, ainsi que leur importance dans le cycle global du soufre, étaient inconnus. Nous démontrons ici que l’enzyme bactérienne MddA, précédemment connue sous le nom de méthanethiol S -méthyltransférase, pourrait méthyler le H 2 S inorganique en DMS. Nous déterminons les résidus clés impliqués dans la catalyse MddA et proposons le mécanisme de méthylation du H 2 S S. Ces résultats ont permis l’identification ultérieure d’enzymes MddA fonctionnelles dans d’abondantes haloarchées et dans une gamme diversifiée d’algues, élargissant ainsi l’importance de la méthylation de H 2 S médiée par MddA à d’autres domaines de la vie. De plus, nous apportons la preuve que la méthylation du H 2 S, S constitue une stratégie de détoxification chez les micro-organismes. Le gène mddA était abondant dans divers environnements, notamment les sédiments marins, les sédiments lacustres, les sources hydrothermales et les sols. Ainsi, l’importance de la méthylation du H 2 S inorganique induite par MddA pour la production mondiale de DMS et le cycle du soufre a probablement été considérablement sous-estimée.
Le sulfate stimule différemment mais n’est pas respiré par diverses archées méthanotrophes anaérobies
https://www.nature.com/articles/s41396-021-01047-0 [51]
L’oxydation anaérobie du méthane (AOM) couplée aux sulfates est un puits de méthane majeur dans les sédiments marins. Plusieurs lignées d’archées anaérobies méthanotrophes (ANME) coexistent souvent dans les sédiments et catalysent ce processus de manière syntrophique avec les bactéries sulfato-réductrices (SRB), mais les différences potentielles dans l’écophysiologie de l’ANME et les mécanismes de syntrophie restent non résolues. Un analogue de l’acide humique, l’anthraquinone 2,6-disulfonate (AQDS), pourrait découpler la méthanotrophie archée de la réduction bactérienne du sulfate et servir d’accepteur terminal d’électrons pour l’AOM (AOM couplée à l’AQDS). Le soufre fait partie intégrante des consortiums AOM pour plusieurs raisons. En tant qu’accepteur d’électrons terminal prédominant pour l’AOM dans les environnements marins, on pense généralement que le sulfate est réduit par les partenaires SRB.
Au-delà de la respiration microbienne, le soufre est un élément essentiel dans tous les organismes, représentant environ 1 % de la biomasse d’une cellule et joue un rôle clé dans de nombreux processus anabolisants en tant que composant des acides aminés et des coenzymes avec des états d’oxydation du soufre variant de +4 à -2. De plus, des composés organosulfurés sont également produits en tant que métabolites secondaires et ont été signalés par les consortiums. L’oxydation du méthane a été couplée à la réduction des sulfates avec une stœchiométrie proche du rapport 1:1 attendu.
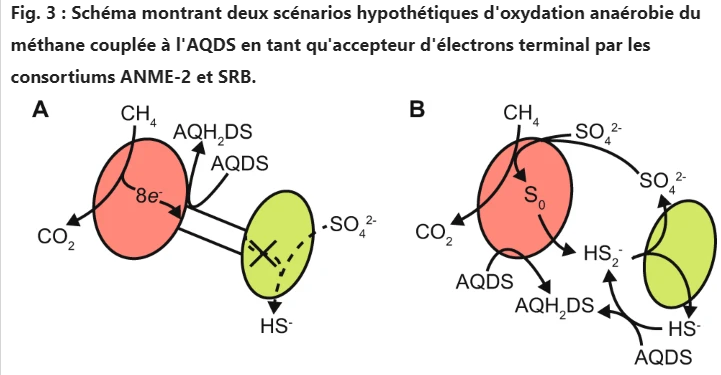
Alors que les membres de l’ANME-2a et de l’ANME-2c codent tous deux pour des homologues des étapes en aval de la voie de réduction assimilatoire des sulfates, l’absence de mécanisme pour former l’adénosine 5′-phosphosulfate indique que la stimulation du sulfate par l’ANME- 2a ne se produit pas par assimilation canonique du sulfate. Alternativement, le sulfate pourrait être impliqué dans l’anabolisme de l’ANME-2 sans être réduit. Par exemple, il est possible que le sulfate soit utilisé par l’ANME-2 pour produire un composé actif rédox, comme le sulfate de thioquinoxalinol identifié dans les consortiums ANME-1a. Lorsqu’un groupe ester sulfate est ajouté au thioquinoxalinol, le sulfate de thioquinoxalinol résultant devient plus soluble dans l’eau et donc plus facilement disponible pour la navette rédox. Si une molécule similaire contenant du soufre actif rédox est produite par l’ANME-2 et utilisée pour la navette électronique, cela pourrait conduire à des taux accrus d’AOM couplés à l’AQDS et à une activité anabolique supérieure à celle observée à partir de l’EET seul. Il a été rapporté que l’AQDS et les composés associés inhibent les micro-organismes, y compris les bactéries sulfato-réductrices, probablement en raison de leur similitude structurelle avec les antibiotiques actifs redox une fois transportés dans le cytoplasme cellulaire. Certaines bactéries et archées, en revanche, y compris les membres des Methanosarcinales , non seulement tolèrent l’AQDS, mais respirent directement cet analogue humique actif rédox. Les voies biochimiques sous-jacentes utilisées par l’ANME-2 pour tolérer et réduire l’AQDS sont inconnues, mais des informations peuvent être tirées des mécanismes connus de réduction de l’AQDS chez d’autres micro-organismes.
Le sulfure de diméthyle intervient dans les interactions microbiennes prédateur-proie entre le zooplancton et les algues dans l’océan
https://pubmed.ncbi.nlm.nih.gov/34697459/ [52]
Le phytoplancton est un élément clé des cycles océaniques du carbone et du soufre. Lors des épisodes de prolifération, certaines espèces peuvent émettre de grandes quantités de sulfure de diméthyle volatil (DMS) organosoufré dans l’océan et, par conséquent, dans l’atmosphère, où il peut moduler la formation d’aérosols et affecter le climat. Dans les environnements aquatiques, le DMS joue un rôle important en tant que signal chimique médiateur de diverses interactions trophiques. Pourtant, son rôle dans les interactions microbiennes prédateur-proie reste insaisissable avec des preuves contradictoires quant à son rôle dans la défense chimique des algues ou dans la chimio-attraction des brouteurs vers les cellules des proies. Ici, nous avons étudié le rôle de signalisation du DMS lors des interactions zooplancton-algues par manipulation génétique et biochimique de l’enzyme génératrice de DMS d’algues, la diméthylsulfoniopropionate lyase (DL) dans l’algue formant des efflorescences Emiliania huxleyi. Nous avons inhibé l’activité DL dans les cellules d’E. huxleyi in vivo à l’aide de l’inhibiteur sélectif de DL, le 2-bromo-3- (diméthylsulfonio) -propionate , et avons surexprimé le gène codant pour DL dans la diatomée modèle Thalassiosira pseudonana. Nous avons montré que l’activité des algues DL ne servait pas de défense chimique anti-pâturage mais renforçait paradoxalement la prédation par le brouteur Oxyrrhis marina et d’autres microzooplanctons et mésozooplanctons, notamment les ciliés et les copépodes. La consommation de proies d’algues avec une activité DL induite a également favorisé la croissance d’O. marina. Dans l’ensemble, nos résultats démontrent que le pâturage médié par le DMS peut être écologiquement important et répandu au cours de la dynamique proie-prédateur dans les écosystèmes aquatiques. Le rôle du DMS d’algues révélé ici, agissant comme un signal de consommation pour les brouteurs, soulève des questions fondamentales concernant la rétention de son enzyme biosynthétique à travers l’évolution du phytoplancton dominant formant des proliférations dans l’océan.
Interactions océan-atmosphère dans le cycle biogéochimique mondial du soufre
https://www.sciencedirect.com/science/article/abs/pii/030442039090059L [53]
Le sulfate est absorbé par les algues et les plantes, puis réduit et incorporé aux composés organosoufrés. Les algues marines produisent du propionate de diméthylsulfonium (DMSP), qui a une fonction osmorégulatrice mais peut également être clivé par voie enzymatique pour produire le sulfure de diméthyle volatil (DMS). Les tentatives visant à identifier les variables qui contrôlent la production océanique de DMS ont montré qu’il n’existe pas de relations simples avec la biomasse algale ou la productivité primaire, mais suggèrent que la concentration de DMS dans l’océan est régulée par une interaction complexe de spéciation algale et d’interactions trophiques. Une partie du DMS produit biogéniquement se diffuse dans l’atmosphère, où il est oxydé, principalement en sulfate d’aérosol. La capacité de ces particules d’aérosol à nucléer des gouttelettes nuageuses, et ainsi à influencer la réflectivité et la stabilité des nuages, constitue la base d’une boucle de rétroaction géophysiologique proposée impliquant le phytoplancton, le soufre atmosphérique et le climat.
Le sulfure de carbonyle (COS) est produit photochimiquement à partir de matière organique dissoute dans l’eau de mer. Le mécanisme de cette réaction est encore inconnu. La diffusion du COS de l’océan vers l’atmosphère est une source mondialement importante de ce gaz, qui participe au cycle de l’ozone stratosphérique. Le sulfure d’hydrogène et le disulfure de carbone sont produits à la surface des océans par des processus encore non identifiés, qui semblent liés à l’activité biogénique. Pour ces gaz, les océans constituent une source mineure dans la troposphère.
Diagenèse à l’état non stationnaire du soufre organique et inorganique dans les sédiments lacustres
https://www.sciencedirect.com/science/article/abs/pii/S0016703716304860
Le soufre contrôle le sort de nombreux éléments géochimiques présents dans les sédiments lacustres , notamment le fer, le phosphore et des oligo-éléments importants pour l’environnement. Nous avons mesuré la spéciation du soufre lié aux eaux interstitielles et aux sédiments (sulfate et sulfures aqueux, soufre élémentaire, monosulfure de fer , pyrite, soufre organique) et les variables géochimiques de soutien (carbone, oxygène, fer) dans les sédiments d’un milieu oxygéné en permanence et d’un bassin saisonnièrement anoxique d’un lac oligotrophe au Québec, en utilisant une combinaison d’analyses d’eau interstitielle, d’extractions séquentielles et d’absorption des rayons X près de la structure de bordure. Un modèle diagénétique précoce à l’état non stable a été développé et calibré sur ce vaste ensemble de données pour aider à comprendre les voies et à quantifier les taux de transformations S. Les résultats suggèrent que la principale source de S dans les sédiments est la sédimentation de l’ester-sulfate organique (RO-SO 3 -H). L’hydrolyse de ces composés fournit une source supplémentaire de sulfate pour l’oxydation microbienne anaérobie de la matière organique sédimentaire, libérant du sulfure dans l’eau interstitielle. Les espèces S réduites liées aux solides s’accumulent sous forme de thiols (R-SH) et de sulfures de fer dans le bassin perpétuellement oxygéné et saisonnièrement anoxique, respectivement. La constante de vitesse estimée par le modèle pour la formation de R-SH est inférieure à celle estimée précédemment pour ce site lacustre particulier, mais similaire à celle proposée pour les sédiments du plateau marin . Les profils S des sédiments solides portent cependant l’empreinte de l’apport de sulfate dans le lac en fonction du temps. Les enrichissements en sulfure de fer formés au cours des dernières décennies de dépôts atmosphériques élevés de SO 4 sont actuellement en train de se dissoudre. Dans les sédiments du bassin perpétuellement oxygéné, cette réaction entrave l’accumulation d’hydroxyde de Fe (III) (oxy) près de l’interface sédiment-eau.
La biogéochimie du soufre (S) est complexe et d’un grand intérêt en raison de son impact sur de nombreux processus environnementaux fondamentaux, allant de la productivité primaire terrestre et du métabolisme microbien des sédiments, jusqu’à la matière organique (MO). préservation à long terme. Dans les lacs, le cycle du S dans les sédiments est dynamique spatialement et temporellement, et étroitement couplé à d’autres espèces et processus biogéochimiques, car il participe à un réseau complexe de réactions géochimiques impliquant 8 états de valence, allant de + 6 (par exemple, sulfate, SO 4 2− ), 0 (par exemple, soufre élémentaire, S 0 ), à −2 (par exemple, sulfure, S −II et soufre organique réduit).
La majorité des flux de S vers les lacs proviennent des sols et des zones humides du bassin versant, généralement sous forme de matière organique allochtone contenant du S lié à l’oxygène (par exemple, RO-SO 3 – Groupes H tels que l’ester-SO 4 et d’autres organosulfates) ou S liés au carbone (par exemple, groupes R-SH tels que les acides aminés et autres thiols). Ces deux derniers groupes peuvent constituer jusqu’à 98 % du pool de S du sol. Les flux aqueux de S vers les lacs proviennent de la minéralisation de ces matières organiques (Zhao et al., 2006), de l’altération des roches ou du dépôt atmosphérique direct de SO 4 2−. En entrant dans la colonne d’eau, le S organique (S org ) peut être utilisé par les micro-organismes via des enzymes qui clivent les liaisons RO-SO 3 -H (par exemple, l’aryl sulfatase) et R-SH (par exemple, la cystéine liase), reconstituant ainsi SO 4 2− et HS − , respectivement. Les sources de S à la surface des sédiments comprennent le S dérivé du bassin versant, le SO 4 2− de la colonne d’eau ainsi que l’organisation S se déposant avec le seston. Dans les sédiments, une fois le dioxygène épuisé, le SO 4 2− est principalement réduit en sulfure par réduction dissimilatrice, à partir de laquelle les bactéries sulfato-réductrices gagnent de l’énergie pour leur croissance et leur entretien.
Le sulfure réagit avec Fe pour former du monosulfure amorphe en pyrite cristalline (FeS 2(s) ). La transformation de FeS (m)s en FeS 2(s) est influencée par divers facteurs, notamment le rapport entre Fe réactif et S −II disponible, l’inhibition par carbone organique labile, la capacité du S 0 (aq) à favoriser la pyritisation via la voie des polysulfures et l’effet du pH sur la cinétique de transformation du FeS m(s). Dans l’ensemble, les interactions entre S et Fe dans les eaux anoxiques, les sols gorgés d’eau et les sédiments ont été largement étudiées.
Le sulfure peut également être séquestré sous forme de S org, par exemple sous forme de R-SH lors de la sulfuration de la OM et de la réduction assimilatoire du SO 4 2− . L’incorporation de sulfures dans la MO a été fréquemment rapportée dans les sédiments marins, mais moins dans les lacs malgré des preuves de concentrations élevées de thiols dans les sédiments d’eau douce. Les sulfures peuvent en outre être oxydés pour former des espèces S 0 , abondantes à la fois dans les phases solides et aqueuses. S 0 réagit avec Fe et les métaux (loïdes), et sert de précurseur à la formation de composés S org , y compris des espèces volatiles.
Ici, nous nous appuyons sur deux approches établies pour acquérir une compréhension quantitative du cycle biogéochimique du S dans les sédiments : des mesures détaillées de spéciation en phase solide et la modélisation diagénétique du transport réactif. Le tableau 1 répertorie 14 études qui ont inclus le cycle du S dans des modèles diagénétiques pour les sédiments d’eau douce, dont neuf présentent des profils de profondeur mesurés des espèces de S par rapport auxquels les prédictions du modèle sont évaluées. Dans cinq cas, SO 4 2− est la seule espèce considérée tandis que dans les quatre autres, les données sur les espèces aqueuses réduites de S (c’est-à-dire S −II ) et sur FeS m(s) et FeS 2(S) sont également prises en compte. Aucun des modèles n’a reproduit les caractéristiques des profils de profondeur de S TOT en phase solide, ni n’a été testé par rapport aux mesures d’autres espèces dominantes en phase solide dans les sédiments lacustres, telles que S org, ou contre les mesures de S 0 , omniprésente aussi bien dans les sédiments que dans l’eau interstitielle.
En reconnaissant l’importance cruciale du S dans le contrôle du devenir des nutriments et des éléments majeurs et traces, notre objectif est d’améliorer notre compréhension quantitative de la spéciation et du devenir du S dans les sédiments d’eau douce. En effet, une intégration plus complète du cycle S dans les modèles diagénétiques constitue un tremplin pour une prédiction plus précise des cycles biogéochimiques d’autres éléments, tels que C, O, Fe et P. Nous combinons un vaste ensemble de données sur la spéciation du S en phase aqueuse et solide, avec un modèle de transport réactif mis à jour pour S, C, O et Fe. diagenèse. Nous estimons les taux de transformation du S organique et inorganique dans les sédiments de deux bassins lacustres oligotrophes connaissant des conditions d’oxygénation différentes tout au long de l’année, nous permettant de quantifier les voies de séquestration du S dans des conditions oxiques et anoxiques.
Émissions de composés soufrés biogènes et leur régulation par les nutriments lors d’une prolifération d’Ulva prolifera dans la mer Jaune
https://www.sciencedirect.com/science/article/abs/pii/S0025326X20310031 [54]
La marée verte est un phénomène écologique anormal provoqué par la croissance excessive et l’accumulation d’algues du phylum Chlorophyta. Les plus grandes proliférations de macroalgues au monde causées par Ulva prolifera se produisent chaque été depuis 2007 dans le sud de la mer Jaune. Ces proliférations peuvent entraîner des pertes économiques en menaçant l’aquaculture et le tourisme, en plus des conséquences écologiques résultant des changements dans la structure et la fonction de la communauté marine, qui ont suscité des recherches approfondies.
Les proliférations d’ U. prolifera proviennent généralement des eaux côtières du Jiangsu fin avril ou début mai de chaque année, apparaissant au début sous la forme de petites parcelles d’algues flottantes, mais s’étendant pour couvrir de vastes zones du sud de la mer Jaune lorsque les conditions de croissance sont favorables. Puis, sous l’influence des courants océaniques et de la mousson du sud-est en été, ces algues sont transportées vers le nord. Une quantité importante de macroalgues arrive régulièrement sur la côte de Qingdao vers le mois de juin de chaque année. Finalement, la floraison se dissipe et a disparu fin août. Lorsque la prolifération d’algues vertes se produit, l’environnement chimique de l’eau de mer peut être considérablement modifié. La photosynthèse efficace d’ U. prolifera pendant la période de croissance rapide peut rapidement assimiler le carbone inorganique, ce qui peut provoquer une augmentation brutale du pH. La décomposition d’ U. prolifera peut consommer une grande quantité d’oxygène pendant la période de déclin, ce qui entraîne des conditions anoxiques dans l’eau de mer locale et affecte ainsi la structure de la communauté, la biomasse et les habitants. En outre, les accumulations massives de macroalgues lors d’une prolifération peuvent constituer une source importante de matière organique dissoute via la photosynthèse et d’autres processus métaboliques dans les eaux côtières, ce qui peut avoir un impact significatif sur la composition organique originale de l’eau de mer régionale. Cependant, on ne sait toujours pas comment les eaux côtières et les écosystèmes de Qingdao réagissent à ces marées vertes régulières à grande échelle.
L’ Ulva peut rapidement assimiler et utiliser du sulfate et synthétiser du diméthylsulfoniopropionate (DMSP), ce qui non seulement affecte l’environnement écologique en tant que l’une des sources de soufre et de carbone les plus importantes pour de nombreuses bactéries marines, mais peuvent avoir des effets négatifs sur la croissance d’autres espèces d’algues. La croissance rapide et abondante d’ Ulva en fait un contributeur clé au cycle biogéochimique côtier ; son rôle dans les cycles du soufre marin est particulièrement important car il produit des niveaux élevés de DMSP. Les concentrations de DMSP dans les algues vertes du genre Ulva peuvent atteindre 200 à 300 μmol g −1 de masse sèche. Le DMSP dans l’océan est dégradé par deux voies principales, soit par déméthylation pour produire de l’acide acrylique (AA) et du méthanethiol, soit par dégradation par le DMSP. lyase produite par le phytoplancton et les bactéries qui génère du diméthylsulfure équimolaire (DMS) et de l’AA.
Le DMS est le composé soufré volatil biogénique marin le plus abondant et représente environ 90 % du flux de soufre de l’océan vers l’atmosphère. De nombreux chercheurs ont conclu que les produits d’oxydation du DMS contribuent directement aux noyaux de condensation des nuages (CCN), qui à leur tour peuvent affecter le climat en augmentant l’albédo des aérosols, bien que certains les chercheurs ont remis en question cette affirmation. Parc et coll. (2017) ont constaté que la contribution du SO 4 2− dérivé du DMS au SO 4 2− total de l’aérosol était multipliée par 7 au cours d’une période de prolifération du phytoplancton arctique par rapport à la période de brume précédente.
L’acide acrylique, un acide gras à chaîne courte que l’on trouve couramment dans l’océan, peut être généré par le clivage du DMSP, mais il est également abondant dans les apports terrestres des eaux côtières. Des recherches antérieures ont démontré que les AA sont une source importante de carbone pour les communautés bactériennes , mais qu’ils peuvent, dans des conditions spécifiques, avoir des effets antibactériens.
À ce jour, les changements de DMSP, DMS et AA dans l’eau de mer provoqués par les proliférations d’U. prolifera ne sont toujours pas clairs. On sait peu de choses sur la manière dont le DMS libéré par U. prolifera contribue aux aérosols dans l’atmosphère ou sur la façon dont il affecte l’écologie et le climat dans les régions en floraison . Malheureusement, les émissions de composés soufrés biogènes lors d’une prolifération d’ U. prolifera n’ont pas été entièrement évaluées car le cycle de prolifération n’a pas été étudié dans son intégralité. Les distributions du DMS, du DMSP, de l’AA et de plusieurs autres paramètres associés ont été mesurées, ainsi que les relations entre ces derniers. les variables mesurées ont été évaluées et discutées. De plus, nous avons également examiné comment le changement des concentrations de DMS, DMSP et AA est régulé par la disponibilité des nutriments en utilisant une expérience en mésocosme en même temps que la prolifération d’ U. prolifera . Cette étude contribuera à approfondir notre compréhension de la réponse biogéochimique aux événements de marée verte, avec un accent particulier sur le cycle du soufre, ainsi que sur les effets sur l’environnement écologique et le climat de la mer Jaune.
Pendant la floraison, les concentrations moyennes de DMS et de DMSPt étaient environ 4 et 2 fois supérieures à celles rapportées en août 2006 (5,91 et 41,38 nmol L −1 , respectivement) lorsqu’il n’y avait pas de prolifération d’U. prolifera dans les eaux de surface. Les changements dans les concentrations de DMS, DMSPd, DMSPt et AA dans les eaux côtières ont été étudiés à trois stades différents d’une prolifération d’ U. prolifera au cours de l’été 2018. Les concentrations de DMS, DMSP et AA dans les eaux de surface se sont révélées significativement plus élevées. Des pics de concentrations de DMSP et d’AA ont été observés à la fin de la période de floraison, indiquant qu’une grande quantité de DMS(P) a été libérée pendant le déclin d’ U. prolifera.
Oxydation anaérobie du propane couplée à la réduction des nitrates par une lignée de la classe Symbiobacteriia
https://www.nature.com/articles/s41467-022-33872-y [63]
On pense que les micro-organismes anaérobies jouent un rôle essentiel dans la régulation du flux d’alcanes gazeux à chaîne courte (SCGA) des écosystèmes terrestres et aquatiques vers l’atmosphère. Il a été confirmé que le sulfate agit comme accepteur d’électrons favorisant l’oxydation anaérobie microbienne des SCGA, mais plusieurs autres accepteurs énergétiquement plus favorables coexistent avec ces gaz dans des environnements anaérobies. Ici, nous montrons qu’un bioréacteur ensemencé avec de la biomasse provenant d’une installation de traitement des eaux usées peut effectuer une oxydation anaérobie du propane couplée à une réduction des nitrates en diazote gazeux et en ammonium. Le bioréacteur a fonctionné pendant plus de 1 000 jours et nous avons utilisé des expériences de marquage au 13 C et au 15 N, ainsi que des analyses métagénomiques, métatranscriptomiques, métaprotéomiques et métabolites pour caractériser la communauté microbienne et les processus métaboliques. Les données suggèrent collectivement qu’une espèce représentant un nouvel ordre au sein de la classe bactérienne Symbiobacteriia est responsable de l’oxydation du propane dépendante du nitrate observée. Le génome fermé de cet organisme, que nous désignons sous le nom de « Candidatus Alkanivorans nitratireducens », code les voies d’oxydation du propane en CO 2 via l’ajout de fumarate et de réduction des nitrates, avec tous les gènes clés exprimés lors de l’oxydation du propane dépendante des nitrates. Nos résultats suggèrent que le nitrate est un puits d’électrons pertinent pour l’oxydation du SCGA dans les environnements anaérobies, constituant un nouveau lien à médiation microbienne entre les cycles du carbone et de l’azote.
Une quantité considérable de gaz naturel est générée à partir de sédiments des grands fonds marins et de suintements d’hydrocarbures dans les marges continentales et les écosystèmes terrestres. La majeure partie du gaz naturel rejeté est consommée par des micro-organismes dans les zones anoxiques avant que le gaz ne se diffuse dans les environnements oxiques et dans l’atmosphère. Les études sur l’oxydation anaérobie du gaz naturel se sont concentrées sur le puissant gaz à effet de serre, le méthane, comme composant le plus abondant (~ 60 à 90 %). Cependant, les alcanes gazeux à chaîne courte (SCGA), notamment l’éthane, le propane, le n-butane et l’ isobutane , sont également des constituants importants du gaz naturel (jusqu’à environ 20 %) et sont d’importants précurseurs de l’ozone et des aérosols organiques. Les émissions atmosphériques mondiales de SCGA sont estimées à 9,2 à 9,6 Tg an −1 pour l’éthane, à 9,6 à 10,5 Tg an −1 pour le propane, à 10 Tg an −1 pour le butane et à 4,2 Tg an −1 pour l’iso -butane.
L’oxydation anaérobie du méthane (AOM) a été largement étudiée et est relativement bien comprise. Les archées méthanotrophes anaérobies (ANME) oxydent le méthane via la voie de la méthanogenèse inverse, y compris le complexe clé méthyl-CoM réductase (MCR). L’ANME transporte les électrons du méthane vers les bactéries sulfato-réductrices syntrophiques (SRB) ou directement vers la réduction des nitrates et des oxydes métalliques. La bactérie « Candidatus Méthylomirabilis oxyfera » réalise l’OMA via la voie « intra-aérobie » d’oxydation du méthane en utilisant le nitrite comme seul accepteur d’électrons. Les micro-organismes associent probablement également l’oxydation des SCGA à la réduction de ces accepteurs d’électrons dans des conditions anoxiques . ‘Californie . M. oxyfera ‘ s’est révélé capable d’oxyder l’éthane et le propane via la méthane monooxygénase, bien qu’il reste à démontrer si ces sources de carbone soutiennent la croissance. À ce jour, le sulfate a été le seul puits d’électrons identifié pour soutenir l’oxydation anaérobie des SCGA. L’isolat deltaprotéobactérien Desulfosarcina aeriophaga BuS5 oxyde le propane et le butane via la voie d’addition de fumarate générant des succinates alkyl-substitués, un processus médié par l’alkylsuccinate synthase (ASS) , et réduit directement le sulfate en sulfure. Il a été découvert qu’un enrichissement de l’archéon « Candidatus Syntrophoarchaeum » activait le butane via la formation de butyl-coenzyme M, un processus catalysé par un MCR divergent, avec des équivalents réducteurs canalisés vers des partenaires syntrophiques SRB. De même, l’oxydation anaérobie de l’éthane a été liée au « Candidatus Argoarchaeum ethanivorans », qui codait également pour un complexe de type MCR et il a été suggéré qu’il forme une relation syntrophique avec SRB. Cependant, un archéon capable de médier l’oxydation anaérobie du propane directement couplée à la réduction des sulfates n’a pas encore été identifié.
Comme le sulfate, le nitrate est un accepteur d’électrons courant dans les écosystèmes naturels et est utilisé par l’archéon ANME ‘ Ca’ . Methanoperedens nitroreducens’ pour AOM. La réduction des nitrates couplée à l’oxydation des SCGA, en utilisant le propane comme exemple) est également plus réalisable thermodynamiquement par rapport aux réactions d’oxydation des SCGA couplées à la réduction des sulfates ( △ G o’ = −102 kJ/mol de propane), mais n’a pas encore été lié à une quelconque lignée microbienne.
Méthylation aérobie du sulfure d’hydrogène en sulfure de diméthyle dans divers micro-organismes et environnements
https://www.nature.com/articles/s41396-023-01430-z [64]
Le sulfure de diméthyle (DMS) est la principale source de biosoufre émis dans l’atmosphère et joue un rôle clé dans le cycle mondial du soufre et potentiellement dans la régulation du climat. On pense que le principal précurseur du DMS est le diméthylsulfoniopropionate. Cependant, le sulfure d’hydrogène (H 2 S), un volatile largement répandu et abondant dans les environnements naturels, peut être méthylé en DMS. Les micro-organismes et les enzymes qui convertissent le H 2 S en DMS, ainsi que leur importance dans le cycle global du soufre, étaient inconnus. Nous démontrons ici que l’enzyme bactérienne MddA, précédemment connue sous le nom de méthanethiol S -méthyltransférase, pourrait méthyler le H 2 S inorganique en DMS. Nous déterminons les résidus clés impliqués dans la catalyse MddA et proposons le mécanisme de méthylation du H 2 S. Ces résultats ont permis l’identification ultérieure d’enzymes MddA fonctionnelles dans d’abondantes haloarchées et dans une gamme diversifiée d’algues, élargissant ainsi l’importance de la méthylation de H 2 S médiée par MddA à d’autres domaines de la vie. De plus, nous apportons la preuve que la méthylation du H 2 S S constitue une stratégie de détoxification chez les micro-organismes. Le gène mddA était abondant dans divers environnements, notamment les sédiments marins, les sédiments lacustres, les sources hydrothermales et les sols. Ainsi, l’importance de la méthylation du H 2 S inorganique induite par MddA pour la production mondiale de DMS et le cycle du soufre a probablement été considérablement sous-estimée.
Le sulfure de diméthyle intervient dans les interactions microbiennes prédateur-proie entre le zooplancton et les algues dans l’océan
https://pubmed.ncbi.nlm.nih.gov/34697459/ [55]
Le phytoplancton est un élément clé des cycles océaniques du carbone et du soufre. Lors des épisodes de prolifération, certaines espèces peuvent émettre de grandes quantités de sulfure de diméthyle volatil (DMS) organosoufré dans l’océan et, par conséquent, dans l’atmosphère, où il peut moduler la formation d’aérosols et affecter le climat. Dans les environnements aquatiques, le DMS joue un rôle important en tant que signal chimique médiateur de diverses interactions trophiques. Pourtant, son rôle dans les interactions microbiennes prédateur-proie reste insaisissable avec des preuves contradictoires quant à son rôle dans la défense chimique des algues ou dans la chimio-attraction des brouteurs vers les cellules des proies. Ici, nous avons étudié le rôle de signalisation du DMS lors des interactions zooplancton-algues par manipulation génétique et biochimique de l’enzyme génératrice de DMS d’algues, la diméthylsulfoniopropionate lyase (DL) dans l’algue formant des efflorescences Emiliania huxleyi. Nous avons inhibé l’activité DL dans les cellules d’E. huxleyi in vivo à l’aide de l’inhibiteur sélectif de DL, le 2-bromo-3- (diméthylsulfonio) -propionate, et avons surexprimé le gène codant pour DL dans la diatomée modèle Thalassiosira pseudonana. Nous avons montré que l’activité des algues DL ne servait pas de défense chimique anti-pâturage mais renforçait paradoxalement la prédation par le brouteur Oxyrrhis marina et d’autres microzooplanctons et mésozooplanctons, notamment les ciliés et les copépodes. La consommation de proies d’algues avec une activité DL induite a également favorisé la croissance d’O. marina. Dans l’ensemble, nos résultats démontrent que le pâturage médié par le DMS peut être écologiquement important et répandu au cours de la dynamique proie-prédateur dans les écosystèmes aquatiques. Le rôle du DMS d’algues révélé ici, agissant comme un signal de consommation pour les brouteurs, soulève des questions fondamentales concernant la rétention de son enzyme biosynthétique à travers l’évolution du phytoplancton dominant formant des proliférations dans l’océan.
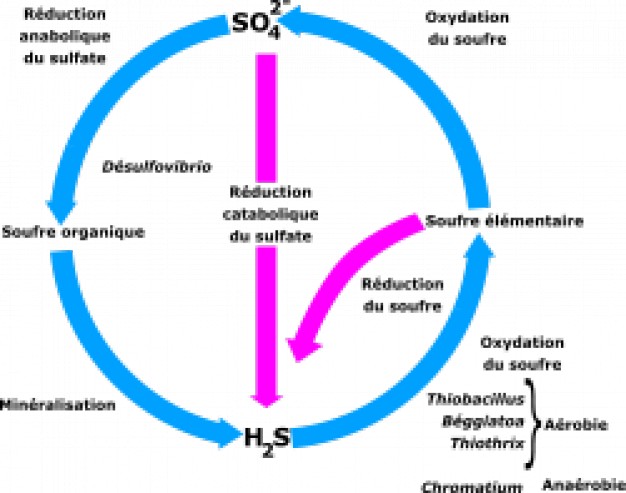
Modélisation de la diffusion du sulfure de diméthyle dans la couche limite externe des algues : implications pour les rôles mutualistes et de signalisation
https://pubmed.ncbi.nlm.nih.gov/30246477/ [55]
Le sulfure de diméthyle (DMS), une espèce de soufre organique dominante à la surface des océans, peut agir comme une molécule de signalisation et contribuer aux interactions mutualistes entre les bactéries et les algues marines. Ces fonctions proposées dépendent de la concentration de DMS au voisinage des micro-organismes. Ici, nous avons modélisé l’enrichissement en DMS à la surface des cellules d’algues marines libérant du DMS en fonction du taux de production de DMS, du rayon des cellules d’algues et de la turbulence. Nos résultats montrent que la concentration de DMS à la surface du phytoplancton non stressé avec de faibles taux de production de DMS peut être enrichie de <1 nM, alors que pour les algues soumises à un stress mécanique avec des activités élevées de l’enzyme DMSP-lyase (un coccolithophore et un dinoflagellé), la surface cellulaire du DMS les enrichissements peuvent atteindre environ 10 nM et pourraient potentiellement atteindre des niveaux de µM dans les grandes cellules. Ces enrichissements en DMS sont bien supérieurs à la concentration médiane de DMS à la surface de l’océan (1,9 nM) et peuvent donc attirer et soutenir la croissance des bactéries vivant dans la phycosphère. Les bactéries, à leur tour, peuvent fournir des chélateurs photoactifs du fer (sidérophores) qui améliorent l’absorption du fer par les algues et fournissent des facteurs de croissance aux algues tels que les auxines et les vitamines. La présente étude met en évidence de nouvelles connaissances sur l’étendue et l’impact des enrichissements en DMS à micro-échelle sur les surfaces d’algues, contribuant ainsi à notre compréhension des rôles potentiels chimioattractants et mutualistes du DMS dans les micro-organismes marins.
Production biogénique de DMSP et sa dégradation en DMS – leurs rôles dans le cycle global du soufre
https://pubmed.ncbi.nlm.nih.gov/31231779/ [56]
Le sulfure de diméthyle (DMS) est la forme de soufre volatil la plus abondante dans les océans de la Terre et est principalement produit par le clivage enzymatique du diméthylsulfoniopropionate (DMSP). Le DMS et le DMSP jouent un rôle important dans le cycle mondial du soufre et peuvent affecter le climat. Le DMSP est proposé pour servir d’osmolyte, de dissuasion du broutage, de molécule de signalisation, d’antioxydant, de cryoprotecteur et/ou de puits pour l’excès de soufre. On a longtemps cru que seuls les eucaryotes marins comme le phytoplancton produisaient du DMSP. Cependant, nous avons récemment découvert que les bactéries marines hétérotrophes peuvent également produire du DMSP, ce qui en fait une source potentiellement importante de DMSP. À l’heure actuelle, une enzyme de synthèse de DMSP procaryote et deux eucaryotes ont été identifiées. Les bactéries marines hétérotrophes sont probablement les principaux dégradants du DMSP, en utilisant deux voies connues : la déméthylation et le clivage. De nombreux phytoplanctons et certains champignons peuvent également cliver le DMSP. Jusqu’à présent, sept DMSP lyases procaryotes et une eucaryotes différentes ont été identifiées. Cette revue décrit le modèle de distribution mondiale du DMSP et du DMS, les gènes connus pour la biosynthèse et le clivage du DMSP, ainsi que les fonctions physiologiques et écologiques de ces molécules organosulfurées importantes, ce qui améliorera la compréhension des mécanismes de production du DMSP et du DMS et de leurs rôles dans l’environnement.
Aperçus récents sur la biosynthèse et le catabolisme du diméthylsulfoniopropionate océanique
https://pubmed.ncbi.nlm.nih.gov/35611751/ [57]
Le diméthylsulfoniopropionate (DMSP), un composé organosoufré d’importance mondiale, est produit en quantités prodigieuses (2,0 Pg de soufre) chaque année dans le milieu marin par le phytoplancton, les macroalgues, les bactéries hétérotrophes, certains coraux et certaines plantes supérieures. Il s’agit d’un osmolyte marin important et d’une molécule précurseur majeure pour la production de sulfure de diméthyle (DMS), un gaz volatil actif sur le plan climatique. La synthèse du DMSP s’effectue via trois voies : une « voie » de transamination chez certaines bactéries et algues marines, une « voie » de Met-méthylation chez les angiospermes et les bactéries et une « voie » de décarboxylation chez le dinoflagellé Crypthecodinium. Les enzymes DSYB et TpMMT sont impliquées dans la biosynthèse du DMSP chez les eucaryotes tandis que les bactéries hétérotrophes marines engagent des enzymes clés telles que DsyB et MmtN. Plusieurs communautés bactériennes marines importent du DMSP et le dégradent via des voies de clivage, de déméthylation ou d’oxydation, générant ainsi respectivement du DMS, du méthanethiol et du propionate de diméthylsulfoxonium. Le DMSP est clivé par diverses enzymes DMSP lyase dans les bactéries et via l’enzyme Alma1 dans le phytoplancton. La voie de déméthylation implique quatre enzymes différentes, à savoir DmdA, DmdB, DmdC et DmdD/AcuH. Cependant, les enzymes impliquées dans la voie d’oxydation n’ont pas encore été identifiées. Nous avons passé en revue les progrès récents sur la synthèse et le catabolisme du DMSP et des enzymes impliquées dans ces processus.
L’analyse structure-fonction indique qu’une molécule d’eau en site actif participe au clivage du diméthylsulfoniopropionate par DddK
https://pubmed.ncbi.nlm.nih.gov/30770407/ [58]
L’osmolyte diméthylsulfoniopropionate (DMSP) est produit en quantités pétagrammes dans les environnements marins et joue un rôle important dans le cycle mondial du soufre et du carbone. De nombreux micro-organismes marins catabolisent le DMSP via des DMSP lyases, générant ainsi du sulfure de diméthyle (DMS), un gaz actif pour le climat. Les produits d’oxydation du DMS participent à la formation des noyaux de condensation des nuages et peuvent ainsi influencer les conditions météorologiques et le climat. Les bactéries SAR11 sont les bactéries hétérotrophes marines les plus abondantes ; beaucoup d’entre eux contiennent la DMSP lyase DddK, et leurs transcriptions dddK sont relativement abondantes dans l’eau de mer. Dans un mécanisme catalytique récemment décrit pour DddK, Tyr64 devrait agir comme base catalytique initiant la réaction de β-élimination du DMSP. Il a été proposé que Tyr64 soit déprotoné par coordination avec le cofacteur métallique ou son His96 voisin. Pour approfondir ce mécanisme, nous avons purifié et caractérisé la protéine DddK de la souche HTCC1062 de Pelagibacter ubique et déterminé les structures cristallines de DddK de type sauvage et de ses mutants Y64A et Y122A (portant un changement de Y en A en position 64 ou 122, respectivement) , où le mutant Y122A est complexé avec le DMSP. Les analyses structurales et mutationnelles soutiennent largement le rôle catalytique de Tyr64, mais pas la méthode de sa déprotonation. Nos données indiquent qu’une molécule d’eau active dans le site actif de DddK joue un rôle important dans la déprotonation de Tyr64 et que cela est bien plus probable que la coordination avec le métal ou His96. L’alignement des séquences et l’analyse phylogénétique suggèrent que le mécanisme catalytique proposé de DddK a une signification universelle. Nos résultats fournissent de nouvelles informations mécanistiques sur DddK et enrichissent notre compréhension de la génération de DMS par les bactéries SAR11. IMPORTANCE Le sulfure de diméthyle (DMS), un gaz actif pour le climat, joue un rôle important dans le cycle mondial du soufre et dans la chimie atmosphérique. Le DMS est principalement produit par le clivage bactérien du diméthylsulfoniopropionate marin (DMSP). Lorsqu’il est rejeté dans l’atmosphère depuis les océans, le DMS peut être oxydé photochimiquement en DMSO ou en aérosols sulfates, qui forment des noyaux de condensation des nuages qui influencent la réflectivité des nuages et, par conséquent, la température mondiale. Les bactéries SAR11 sont les bactéries hétérotrophes marines les plus abondantes, et beaucoup d’entre elles contiennent de la DMSP lyase DddK pour cliver le DMSP, générant ainsi du DMS. Dans cette étude, basée sur des analyses structurelles et des tests mutationnels, nous avons révélé le mécanisme catalytique de DddK, qui a une signification universelle chez les bactéries SAR11. Cette étude fournit de nouvelles informations sur le mécanisme catalytique de DddK, conduisant à une meilleure compréhension de la manière dont les bactéries SAR11 génèrent du DMS.
Le stress oxydatif régule un interrupteur métabolique crucial dans la dégradation du diméthylsulfoniopropionate par la bactérie marine Ruegeria pomeroyi
https://pubmed.ncbi.nlm.nih.gov/36301115/ [59]
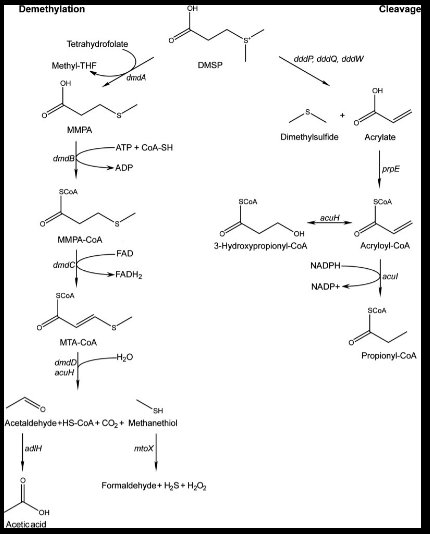
Le diméthylsulfoniopropionate (DMSP) est un composé organique abondant dans les eaux de surface marines et une source de sulfure de diméthyle (DMS), la plus grande source naturelle de soufre dans la haute atmosphère. Les bactéries marines minéralisent le DMSP par la voie de déméthylation ou le transforment en DMS par la voie de clivage. Les facteurs qui déterminent quelle voie est utilisée ne sont pas entièrement compris. Dans des expériences de chémostat, la Roseobacter marine Ruegeria pomeroyi DSS-3 a été exposée à un stress oxydatif soit pendant sa croissance avec H 2 O 2 , soit par mutation du gène codant pour la catalase. Le stress oxydatif a réduit l’expression des gènes dans la voie de déméthylation et a augmenté l’expression de ceux codant pour la voie de clivage. Ces résultats sont contraires à l’hypothèse de la demande en soufre, selon laquelle le métabolisme du DMSP dépend des besoins en soufre des cellules bactériennes. Au lieu de cela, nous trouvons des preuves solides cohérentes avec le contrôle du stress oxydatif sur le passage du métabolisme du DMSP de la déméthylation à la production de DMS chez une bactérie marine écologiquement pertinente. IMPORTANCE Le diméthylsulfoniopropionate (DMSP) est le composé organique de faible poids moléculaire le plus abondant dans les eaux de surface marines et la source de sulfure de diméthyle (DMS), un gaz climatiquement actif qui relie les cycles du soufre marin et terrestre. Les bactéries marines sont les principales consommatrices de DMSP, soit en générant du DMS, soit en consommant du DMSP comme source de carbone et de soufre réduits. Cependant, les facteurs régulant le catabolisme du DMSP chez les bactéries ne sont pas bien compris. Les bactéries marines sont également exposées au stress oxydatif. Des expériences de séquençage d’ARN (RNA-seq) ont montré que le stress oxydatif induit en laboratoire réduisait l’expression des gènes codant pour la consommation de DMSP via la voie de déméthylation et augmentait l’expression des gènes codant pour la production de DMS via la voie de clivage dans la bactérie marine Ruegeria pomeroyi. Ces résultats soutiennent un modèle dans lequel la production de DMS dans l’océan est régulée en partie par le stress oxydatif.
Une nouvelle diméthylsulfoniopropionate lyase dépendant de l’ATP chez des bactéries qui libère du sulfure de diméthyle et de l’acryloyl-CoA
https://pubmed.ncbi.nlm.nih.gov/33970104/
Le diméthylsulfoniopropionate (DMSP) est une molécule organosoufrée abondante et omniprésente dans les environnements marins, jouant un rôle important dans le cycle mondial du soufre et des nutriments. Diverses lyases DMSP présentes dans certaines algues, bactéries et champignons clivent le DMSP pour produire du sulfure de diméthyle gazeux (DMS), un infochimique jouant un rôle important dans la chimie atmosphérique. Ici, nous avons identifié une nouvelle DMSP lyase dépendante de l’ATP, DddX. DddX appartient à la superfamille des acyl-CoA synthétases et se distingue des huit autres DMSP lyases connues. DddX catalyse la conversion du DMSP en DMS via une réaction en deux étapes : la ligature du DMSP avec CoA pour former l’intermédiaire DMSP-CoA, qui est ensuite clivé en DMS et acryloyl-CoA. Le nouveau mécanisme catalytique a été élucidé par des analyses structurelles et biochimiques. DddX se trouve dans plusieurs alphaprotéobactéries, gammaprotéobactéries et Firmicutes, ce qui suggère que cette nouvelle DMSP lyase pourrait jouer un rôle négligé dans les cycles DMSP/DMS.
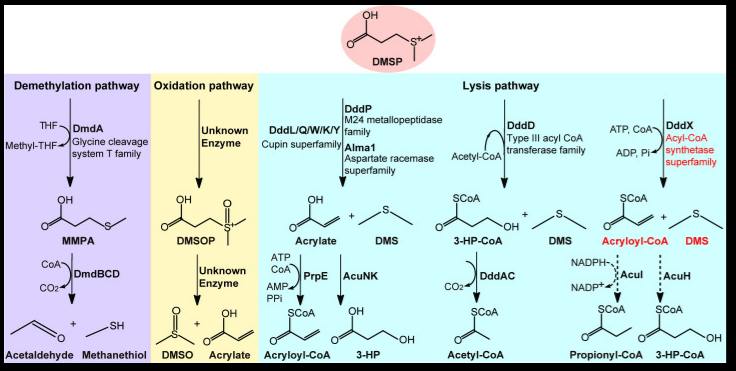
Le cycle mondial du soufre est un ensemble de processus géologiques et biologiques qui font circuler des composés contenant du soufre dans les océans, les roches et l’atmosphère. Le soufre lui-même est essentiel à la vie et important pour la croissance des plantes, d’où son utilisation répandue dans les engrais. Les organismes marins tels que les bactéries, les algues et le phytoplancton produisent en quantités massives un composé soufré particulier, appelé diméthylsulfoniopropionate, ou DMSP. Le DMSP produit dans les océans est facilement converti en un gaz appelé sulfure de diméthyle (DMS), qui constitue la plus grande source naturelle de soufre entrant dans l’atmosphère. Dans l’air, le DMS est converti en sulfate et en d’autres sous-produits qui peuvent agir comme des noyaux de condensation des nuages qui, comme leur nom l’indique, sont impliqués dans la formation des nuages. De cette manière, le DMS peut influencer les conditions météorologiques et le climat, c’est pourquoi il est souvent qualifié de gaz « actif sur le plan climatique ». On sait qu’au moins huit enzymes scindent le DMSP en gaz DMS avec quelques sous-produits. Ces enzymes se trouvent dans les algues, les bactéries et les champignons et sont appelées lyases, en raison de la façon dont elles décomposent leurs composés cibles (DMSP, dans ce cas). Récemment, des chercheurs ont identifié certaines bactéries qui produisent du DMS à partir du DMSP sans utiliser de DMSP lyases connues. Cela suggère qu’il existe d’autres enzymes non identifiées qui agissent sur le DMSP dans la nature et contribuent probablement au cycle mondial du soufre. Li, Wang et coll. a entrepris de découvrir de nouvelles enzymes responsables de la conversion du DMSP produit par les bactéries marines en DMS gazeux. Une nouvelle enzyme appelée DddX a été identifiée et s’est avérée appartenir à une superfamille d’enzymes assez distincte des autres DMSP lyases connues. Li, Wang et coll. a également montré comment DddX pilote la conversion du DMSP en DMS dans une réaction en deux étapes, et que l’enzyme est présente dans plusieurs classes de bactéries. D’autres expériences visant à caractériser la structure protéique de DddX ont également révélé le mécanisme moléculaire de son action catalytique. Cette étude offre des informations importantes sur la manière dont les bactéries marines génèrent le gaz DMS, important sur le plan climatique, à partir du DMSP, conduisant à une meilleure compréhension du cycle mondial du soufre. Il donne aux écologistes microbiens une perspective plus complète de ces processus environnementaux et fournit aux biochimistes des données sur une famille d’enzymes jusqu’alors inconnues pour agir sur des composés contenant du soufre.
Le sulfate stimule différemment mais n’est pas respiré par diverses archées méthanotrophes anaérobies
https://www.nature.com/articles/s41396-021-01047-0 [65]
L’oxydation anaérobie du méthane (AOM) couplée aux sulfates est un puits de méthane majeur dans les sédiments marins. Plusieurs lignées d’archées anaérobies méthanotrophes (ANME) coexistent souvent dans les sédiments et catalysent ce processus de manière syntrophique avec les bactéries sulfato-réductrices (SRB), mais les différences potentielles dans l’écophysiologie de l’ANME et les mécanismes de syntrophie restent non résolues. Un analogue de l’acide humique, l’anthraquinone 2,6-disulfonate (AQDS), pourrait découpler la méthanotrophie archée de la réduction bactérienne du sulfate et servir d’accepteur terminal d’électrons pour l’AOM (AOM couplée à l’AQDS). Ici, dans des expériences en microcosme de sédiments, nous avons examiné les variations de la réponse physiologique entre deux familles d’ANME-2 concomitantes (ANME-2a et ANME-2c) et testé l’hypothèse de la respiration sulfate par ANME-2. Des concentrations de sulfate aussi faibles que 100 µM ont augmenté l’AOM couplée à l’AQDS de près de 2 fois, correspondant aux taux d’AOM couplées au sulfate. Cependant, les partenaires SRB sont restés inactifs dans les microcosmes contenant du sulfate et de l’AQDS et aucune des familles ANME-2 n’a respiré le sulfate, comme le montrent leurs teneurs en soufre cellulaire et leurs activités anabolisantes mesurées par spectrométrie de masse d’ions secondaires à l’échelle nanométrique. L’activité anabolique de l’ANME-2a était significativement supérieure à celle de l’ANME-2c, ce qui suggère que l’ANME-2a était principalement responsable de la stimulation par le sulfate observée de l’AOM couplée à l’AQDS. La transcriptomique comparative a montré une régulation positive significative des transcrits ANME-2a liés à plusieurs transporteurs ABC et une régulation négative du métabolisme central du carbone au cours de l’AOM couplée à l’AQDS par rapport à l’AOM couplée au sulfate. Étonnamment, les gènes impliqués dans l’anabolisme du soufre n’étaient pas exprimés de manière différentielle au cours de l’AOM couplée à l’AQDS avec et sans amendement sulfate. Collectivement, ces données indiquent que les archées ANME-2 sont incapables de respirer du sulfate, mais la disponibilité du sulfate stimule différemment la croissance et l’activité AOM de différentes lignées ANME. L’oxydation anaérobie du méthane (AOM) couplée au sulfate est un processus microbien qui consomme >90 % du flux annuel de méthane provenant des sédiments marins. Une symbiose entre les archées méthanotrophes anaérobies (ANME) et les bactéries sulfato-réductrices (SRB) catalyse l’AOM couplée au sulfate. Depuis la découverte de ces consortiums il y a vingt ans, diverses lignées ANME (par exemple ANME-1a/1b/2a/2b/2c/3) ont été reconnues pour former des partenariats avec différentes lignées SRB (par exemple HotSeep-1, SEEP-SRB1a/1g). ). Ces lignées représentent différentes espèces, genres, familles et même ordres, et codent pour un pangénome étendu. Cette diversité phylogénétique et génomique favorise probablement les différences dans les propriétés métaboliques, les interactions symbiotiques et l’adaptation écologique entre les consortiums AOM dans les écosystèmes de suintement de méthane.
Interactions océan-atmosphère dans le cycle biogéochimique mondial du soufre
https://www.sciencedirect.com/science/article/abs/pii/030442039090059L [61]
Le sulfate est absorbé par les algues et les plantes, puis réduit et incorporé aux composés organosoufrés. Les algues marines produisent du propionate de diméthylsulfonium (DMSP), qui a une fonction osmorégulatrice mais peut également être clivé par voie enzymatique pour produire le sulfure de diméthyle volatil (DMS). Les tentatives visant à identifier les variables qui contrôlent la production océanique de DMS ont montré qu’il n’existe pas de relations simples avec la biomasse algale ou la productivité primaire, mais suggèrent que la concentration de DMS dans l’océan est régulée par une interaction complexe de spéciation algale et d’interactions trophiques. Une partie du DMS produit biogéniquement se diffuse dans l’atmosphère, où il est oxydé, principalement en sulfate d’aérosol. La capacité de ces particules d’aérosol à nucléer des gouttelettes nuageuses, et ainsi à influencer la réflectivité et la stabilité des nuages, constitue la base d’une boucle de rétroaction géophysiologique proposée impliquant le phytoplancton, le soufre atmosphérique et le climat. Le sulfure de carbonyle (COS) est produit photochimiquement à partir de matière organique dissoute dans l’eau de mer. Le mécanisme de cette réaction est encore inconnu. La diffusion du COS de l’océan vers l’atmosphère est une source mondialement importante de ce gaz, qui participe au cycle de l’ozone stratosphérique. Le sulfure d’hydrogène et le disulfure de carbone sont produits à la surface des océans par des processus encore non identifiés, qui semblent liés à l’activité biogénique. Pour ces gaz, les océans constituent une source mineure dans la troposphère.
Production biologique et variation spatiale des composés soufrés diméthylés et leur relation avec le plancton dans le nord de la mer Jaune
https://www.sciencedirect.com/science/article/abs/pii/S0278434315000941 [62]
Dans la couche superficielle de l’océan, le sulfure de diméthyle (DMS) produit par le plancton est le gaz de soufre actif réduit le plus important échangé de la mer vers l’air. Dans l’atmosphère, le DMS peut être oxydé en sulfate non marin et en acide méthane sulfonique, responsable d’une partie de la formation de noyaux de condensation des nuages. Ces processus affectent ainsi l’équilibre radiatif de la Terre à la fois directement, par la rétrodiffusion du rayonnement solaire, et indirectement, par la formation de noyaux de condensation des nuages. Ainsi, la production planctonique de DMS et son émission ultime dans l’atmosphère pourraient contrecarrer la tendance au réchauffement climatique ; à son tour, ce changement climatique pourrait avoir un impact sur la biogenèse du DMS dans l’océan. Dans l’océan, le DMS provient d’un soluté compatible, le diméthylsulfoniopropionate ((CH 3 ) 2 S + CH 2 CH 2 COO − ; DMSP). Ce composé tertiaire sulfonium est un osmolyte, antioxydant et cryo-protecteur synthétisé principalement par une grande variété de phytoplancton marin, de macroalgues et de plantes aquatiques submergées. Le DMSP est largement distribué dans la région euphotique des océans à un niveau plus élevé de 10 à 100 nM dans tous les océans. De plus, la production de DMSP est associée à la spéciation des algues, les Dinophyceae et les Prymnesiophyceae étant les principales sources ; par conséquent, la répartition du DMSP est hétérogène et, dans la plupart des cas, n’est pas strictement liée à la biomasse algale totale . Des observations ont également montré une corrélation entre les concentrations de DMSP et la chlorophylle du phytoplancton dans certaines conditions environnementales. De plus, le DMSP est un substrat crucial dans le réseau trophique microbien dans lequel il fournit suffisamment de soufre et de carbone pour satisfaire les exigences de la croissance bactérienne marine par la médiation biologique de DMSP, affectant en outre directement le rendement DMS du DMSP. Ainsi, le DMS et son précurseur DMSP constituent le réservoir majeur de soufre organique dans le milieu marin et la production et la transformation de ces composés constituent une partie importante du cycle du soufre océanique. La libération et la dégradation du DMSP, ainsi que le renouvellement du DMS, sont étroitement liés aux activités du réseau trophique. Les concentrations de DMS sont déterminées par l’équilibre entre ses mécanismes de production et de perte , qui font partie d’un réseau complexe de processus écologiques et biogéochimiques. Les facteurs qui contrôlent la libération du DMS dans l’eau ne sont pas clairement compris, mais les mécanismes possibles sont l’excrétion métabolique accompagnant la croissance du phytoplancton, le broutage du phytoplancton par le zooplancton, la sénescence des microalgues, l’interaction du bactérioplancton et des agents pathogènes viraux. Une fois produit, le DMS est perdu dans l’eau de mer par trois voies principales : l’échange mer-air, la consommation biologique et la transformation photochimique, ces deux derniers processus conduisant à la formation de diméthylsulfoxyde (DMSO). ). Certes, l’interaction intime entre la formation et la dégradation du DMS apporte une contribution considérable à la concentration in situ du DMS ainsi qu’à sa variabilité spatio-temporelle.
https://agupubs.onlinelibrary.wiley.com/doi/10.1002/2017JC013265
https://www.sciencedirect.com/science/article/abs/pii/S1385110100000307
https://www.sciencedirect.com/science/article/abs/pii/S1385110104001091
https://www.sciencedirect.com/science/article/abs/pii/S0304420310001386
Preuve transcriptomique des activités métaboliques polyvalentes des micro-organismes cyclant le mercure dans les tapis microbiens saumâtres
https://www.nature.com/articles/s41522-021-00255-y
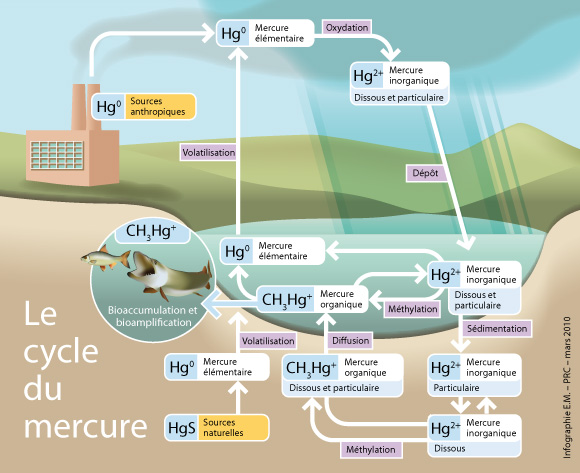
Le méthylmercure, qui se bioamplifie à travers les chaînes alimentaires, est hautement toxique pour la vie aquatique. Sa production et sa dégradation sont largement induites par des transformations microbiennes ; cependant, la diversité et l’activité métabolique des transformateurs du mercure, qui entraînent des concentrations de méthylmercure dans les environnements, restent mal comprises. Les tapis microbiens sont des biofilms épais où coexistent les métabolismes oxiques et anoxiques, offrant ainsi la possibilité d’étudier la complexité des transformations microbiennes du mercure dans des conditions redox contrastées. Ici, nous avons effectué une analyse métagénomique et métatranscriptomique résolue par le génome pour identifier l’activité putative des réducteurs, méthylateurs et déméthylateurs du mercure dans des tapis microbiens fortement contaminés par le mercure. Nos résultats transcriptomiques ont révélé le rôle majeur de micro-organismes rares dans le cycle du mercure. Les méthylateurs de mercure, principalement liés à Desulfobacterota , ont exprimé un large éventail d’activités métaboliques dans les transformations de composés de soufre, de fer, d’azote et d’halogène, étendant les activités connues des méthylateurs de mercure dans des conditions suboxiques à anoxiques. Les processus de détoxification du méthylmercure ont été dissociés dans les tapis microbiens, le clivage du méthylmercure étant effectué par les populations de Thiotrichaceae et de Rhodobacteraceae oxydant les sulfures , tandis que les réducteurs de mercure comprenaient des membres des Verrucomicrobia , Bacteroidetes , Gammaproteobacteria et différentes populations de Rhodobacteraceae . Cependant, la majeure partie de la réduction du mercure a été potentiellement réalisée de manière anaérobie par des Desulfuromonadaceae réducteurs de soufre et de fer , révisant ainsi notre compréhension de l’écophysiologie des transformateurs de mercure.
Impacts de la pollution plastique sur la biogéochimie du carbone marin
https://www.sciencedirect.com/science/article/abs/pii/S0269749120362862
L’un des défis majeurs dans la compréhension de la dynamique de la santé et du fonctionnement des océans est l’impact potentiel de la présence croissante de plastique. Outre leurs effets vérifiés et macroscopiques sur la faune et les habitats marins, les micro et macroplastiques offrent des sites potentiels pour l’activité microbienne et le lessivage chimique . La plupart des plastiques marins se trouvent initialement dans les mètres supérieurs de la colonne d’eau, où les processus biogéochimiques fondamentaux déterminent la productivité marine et la dynamique du réseau trophique. Cependant, des découvertes récentes montrent un continuum d’effets potentiels de ces nouveaux composants marins sur le carbone, les nutriments et les processus microbiens. Dans la présente analyse, nous développons un terrain d’entente entre ces études et identifions les lacunes dans les connaissances sur lesquelles de nouveaux efforts de recherche devraient être concentrés, afin de mieux déterminer les rétroactions potentielles des plastiques sur la biogéochimie du carbone d’un océan en évolution. Une accumulation de plastique à la surface de la mer, un environnement déjà enrichi en matière organique dérivée de l’activité microbienne dans la zone euphotique, favorise en outre la croissance microbienne et la libération de polymères organiques complexes à l’interface air-mer. Ce processus implique 1) une pénétration réduite du rayonnement photosynthétiquement actif (PAR) pour la croissance du phytoplancton et la photosynthèse, 2) une production altérée d’oxygène et de dioxyde de carbone liée à des processus de production/dégradation altérés, et 3) une barrière plus épaisse pour l’air. échange marin de gaz. De plus, le carbone organique dissous (COD) extrait des plastiques augmente l’activité bactérienne hétérotrophe et le remaniement de la matière organique dissoute (MOD), avec pour conséquence une augmentation de la respiration bactérienne et de la consommation d’oxygène. À des profondeurs plus élevées, l’agrégation de la matière organique autour des particules en suspension, comme le plastique, est facilitée par l’interaction du phytoplancton et des bactéries. 4) Le biofilm qui intègre la particule de plastique modifie sa flottabilité et facilite son flux descendant, tout en transférant le carbone organique particulaire (POC) vers l’intérieur de l’océan et éventuellement vers le fond marin. 5) Le biofilm entourant la particule de plastique peut incorporer des nutriments supplémentaires tout au long de son flux descendant, favorisant ainsi une activité microbienne supplémentaire et la libération de composés organiques plus complexes, comme la DOM réfractaire (RDOM), alimentant ainsi un réservoir de carbone réfractaire à long terme en profondeur.
Le plastique peut affecter les organismes marins, du micro au macro, à travers de multiples processus qui impliquent l’ingestion, l’enchevêtrement, la destruction de l’habitat, le transfert de composés toxiques, soit par la libération de composants du plastique lui-même, soit par la concentration et la libération de polluants présents dans l’eau de mer environnante. Des recherches récentes montrent également que les particules de plastique peuvent transporter des espèces pathogènes, conduisant au développement possible d’une résistance aux antibiotiques avec d’autres effets délétères sur la santé.
Tendances mondiales et incertitudes concernant la dénitrification terrestre et les émissions de N 2 O
https://royalsocietypublishing.org/doi/10.1098/rstb.2013.0112 [66]
Le cycle global de l’azote (N) est régi par la fixation de l’azote moléculaire atmosphérique inerte (N 2 ) et la formation de composés azotés réactifs biodisponibles, tels que le nitrate (NO 3 − ), l’ammonium (NH 4 + ) et le N- oxydes. Les humains ont accéléré ce cycle grâce à la production et à l’utilisation d’engrais et à la combustion de combustibles fossiles. La dénitrification est le processus microbien qui élimine le NO 3 − , le réduisant de manière anaérobie en nitrite (NO 2 − ), en oxyde nitrique (NO), en oxyde d’azote (N 2 O), un gaz à effet de serre, et en N 2 . Dans les sols agricoles, le NO 3 − provient principalement des engrais, du fumier animal, des résidus de récolte et de la matière organique du sol. Les substrats organiques doivent d’abord être minéralisés en ammonium par des micro-organismes hétérotrophes, tandis que les bactéries nitrifiantes oxydent ensuite le NH 4 + en NO 2 − et NO 3 − . Dans les écosystèmes naturels, les principales sources d’azote sont la fixation biologique de N 2 et les dépôts atmosphériques. Dans les écosystèmes terrestres, la dénitrification se produit principalement dans les sols, les eaux souterraines et les zones riveraines. D’autres systèmes gérés par l’homme où se produit la dénitrification comprennent les systèmes de stockage du fumier, les usines de traitement des eaux usées, les bioréacteurs et les zones humides construites ou récupérées.
La dénitrification est un processus important car elle entraîne des pertes significatives d’azote dans les systèmes agricoles, tandis qu’en convertissant NO 3 − et NO 2 − en N 2 gazeux , N 2 O et NO x (NO + NO 2 ), la dénitrification réduit le problème de une surcharge globale en N sur le système hydrologique. Ce faisant, la dénitrification transfère cette surcharge dans l’atmosphère, où N 2 O et NO x peuvent avoir des impacts environnementaux négatifs.
La dénitrification étant un processus anaérobie, l’oxygène (O 2 ) est le régulateur le plus important. Le nitrate est l’accepteur d’électrons et la source de N, et le C sert de donneur d’électrons et de source d’énergie pour les bactéries dénitrifiantes hétérotrophes. Le pH du sol a un effet marqué sur la dénitrification, avec des taux plus faibles dans des conditions acides que dans des conditions légèrement alcalines. La dénitrification a une plage de température optimale de 25 à 30 °C. La température contrôle également les taux de décomposition et de nitrification, et régule donc la disponibilité de O 2 , NO 3 − et C organique. La dénitrification du sol ainsi que la production et la consommation de N 2 O sont extrêmement variables dans le temps et dans l’espace en raison de la variabilité de les principaux régulateurs.
Les rejets d’eaux souterraines vers les eaux de surface sont généralement constitués d’un mélange d’eau provenant de différentes parties de l’aquifère, caractérisé par une large gamme de temps de parcours. La concentration de NO 3 − dans les eaux souterraines dépend du temps d’infiltration dans la zone saturée, de la concentration de NO 3 − de cette eau et de la perte par dénitrification lors de son transport. À mesure que l’O 2 devient limitant dans la zone saturée, les micro-organismes hétérotrophes se tournent de plus en plus vers le NO 3 − comme accepteur d’électrons pour l’oxydation du C organique. Ce C organique peut être présent sous forme dissoute ou dans la matrice sédimentaire. Si la pyrite est présente dans les sédiments souterrains, la dénitrification couplée à l’oxydation de la pyrite peut devenir le processus d’élimination dominant. La pyrite est un disulfure de fer commun et se trouve souvent en association avec des dépôts marins non consolidés dans le sous-sol terrestre et en association avec des dépôts de charbon et de schiste.
Les zones riveraines sont des zones humides situées à l’interface entre les composantes terrestres et aquatiques du paysage, qui, de par leur position, contribuent au contrôle des flux de nutriments et d’énergie vers les eaux de surface. La dénitrification riveraine est un mécanisme important pour éliminer le N dans le ruissellement souterrain et les eaux souterraines peu profondes se déplaçant des hautes terres vers les ruisseaux. À l’instar des sols, les émissions de N 2 O des zones riveraines sont causées par un déséquilibre entre la production et la consommation de N 2 O. Ces déséquilibres augmentent à des concentrations élevées d’O 2 et de NO 3 − dans le sol et à un pH faible. Bien que limitées en superficie, les zones riveraines ont été identifiées comme des points chauds pour la dénitrification et l’émission de N 2 O, en particulier le long des cours d’eau d’ordre inférieur.
Les taux de dénitrification et les émissions de N 2 O ont changé à la suite de l’intervention humaine dans le cycle mondial de l’azote. La croissance démographique et le développement économique futurs pourraient conduire à une nouvelle accélération du cycle mondial de l’azote. Quantifier où, quand et dans quelle mesure la dénitrification se produit sur la seule base de mesures est pratiquement impossible. Les modèles sont donc devenus des outils essentiels. Dans cet article, nous abordons le changement possible de la dénitrification terrestre et des émissions de N 2 O des sols, des eaux souterraines et des zones riveraines dans le cadre du changement climatique passé et futur, de l’augmentation de la production alimentaire et énergétique et de l’intensification agricole, à l’aide d’un modèle de simulation. Le terme dénitrification est utilisé pour indiquer la réduction totale des nitrates en N 2 , N 2 O et NO. Des détails sur les données, les modèles et les scénarios utilisés ainsi qu’une partie des résultats sont fournis dans le matériel électronique supplémentaire.
Notre tentative de simuler la dénitrification riveraine et l’émission de N 2 O est la première jamais réalisée et indique que les zones riveraines peuvent être un élément important du paysage pour l’élimination de l’azote au niveau mondial ; étant donné les conditions dans de nombreuses zones riveraines, avec une dénitrification incomplète accompagnée d’une production fractionnée élevée de N 2 O, les émissions mondiales de N 2 O des zones riveraines peuvent dépasser celles des eaux souterraines.
Les scénarios indiquent que l’augmentation progressive du transfert réactif d’azote vers les eaux de surface et l’atmosphère se poursuivra au cours des prochaines décennies, à moins que des améliorations drastiques de la gestion agricole ne soient apportées. Cela peut avoir des conséquences importantes sur la biogéochimie des réseaux fluviaux et des mers côtières, induisant une série de processus, tels que la stimulation de la croissance des plantes, la prolifération d’algues, la décomposition, l’enfouissement et l’hypoxie.
Appauvrissement de la couche d’ozone stratosphérique dû au protoxyde d’azote : influences d’autres gaz
https://royalsocietypublishing.org/doi/abs/10.1098/rstb.2011.0377?intcmp=trendmd [67]
Les effets des émissions anthropiques d’oxyde d’azote (N 2 O), de dioxyde de carbone (CO 2 ), de méthane (CH 4 ) et d’halocarbures sur l’ozone stratosphérique (O 3 ) au cours des XXe et XXIe siècles sont isolés à l’aide d’un modèle chimique. de la stratosphère. L’évolution future de l’ozone dépendra de chacun de ces gaz, le N 2 O et le CO 2 jouant probablement les rôles dominants à mesure que les halocarbures reviennent vers les niveaux préindustriels. Il existe des interactions non linéaires entre ces gaz qui empêchent de séparer sans ambiguïté leurs effets sur l’ozone. Par exemple, l’augmentation du CH 4 au cours du XXe siècle a réduit les pertes d’ozone dues à l’augmentation des halocarbures, et la destruction chimique de l’O 3 par le N 2 O est amortie par les effets thermiques du CO 2 dans la stratosphère moyenne (d’environ 20 % pour le GIEC). Scénario A1B/OMM A1 sur la période 1900-2100). Néanmoins, on s’attend à ce que le N 2 O continue d’être la plus grande émission anthropique d’un composé destructeur d’O 3 dans un avenir prévisible. Les réductions des émissions anthropiques de N 2 O offrent de plus grandes possibilités de réduction de l’épuisement futur de l’O 3 que n’importe quelle autre émission non contrôlée d’halocarbures. Il est également démontré que les niveaux d’O 3 de 1980 étaient affectés par les halocarbures, le N 2 O, le CO 2 et le CH 4 et qu’ils ne constituent donc peut-être pas un bon choix de référence pour la récupération de l’O 3 .
De nombreux processus constituant les cycles globaux des éléments peuvent se produire sans catalyse microbienne, bien que souvent à un rythme considérablement plus faible ; le cycle de l’azote est cependant unique en ce sens qu’il dépend presque exclusivement de réactions redox régulées par des micro-organismes. Par conséquent, les microbes sont non seulement essentiels au maintien du cycle, mais également à la biodisponibilité de l’azote et constituent donc un facteur de contrôle clé de la production primaire. L’importance de la transformation de l’azote dans les sédiments à forte charge organique est éclipsée par d’autres réactions redox telles que la réduction des sulfates. Le déséquilibre des activités métaboliques des nitrifiants et des dénitrifiants maintient des concentrations élevées de nitrate dans toute la colonne de sédiments et est probablement dû à une disponibilité limitée de carbone pour les dénitrifiants.
L’abondance et la diversité des nitrifiants culminent dans les zones de transition rédox profondes
https://www.nature.com/articles/s41598-019-44585-6 [68]
Plus de la moitié des fonds océaniques mondiaux sont recouverts de sédiments pauvres en nutriments, caractérisés par une pénétration profonde de l’oxygène et une prédominance d’azote oxydé. Malgré une faible disponibilité énergétique, cet habitat héberge une vaste population microbienne et les caractéristiques géochimiques suggèrent que les composés azotés sont une source d’énergie essentielle au maintien de cette biomasse. Cependant, les taux métaboliques de transformation de l’azote et leur lien avec la survie microbienne dans cet écosystème à l’échelle mondiale restent pratiquement inconnus. Nous fournissons ici des contraintes quantitatives sur le cycle microbien de l’azote dans les sédiments oligotrophes des océans, du fond marin au sous-sol, sur environ 8 millions d’années. Nous constatons une transformation microbienne active de l’azote dans toute la colonne de sédiments, mais à des taux très faibles. Des pics locaux de diversité et d’abondance de nitrifiants et de dénitrifiants se produisent dans les zones de transition rédox situées profondément dans les sédiments, ce qui indique fortement que ces microbes sont réanimés de leur état de maintenance et recommencent à croître après des millions d’années d’attrition. Les communautés microbiennes présentes dans les sédiments marins ont un impact sur tous les principaux cycles biogéochimiques mondiaux, notamment ceux de l’oxygène, du carbone, de l’azote, du manganèse, du fer et du soufre. En assurant la médiation du flux de composés chimiques entrant et sortant des sédiments, cette biosphère cachée influence la composition de l’océan et de l’atmosphère. De nombreux processus constituant les cycles globaux des éléments peuvent se produire sans catalyse microbienne, bien que souvent à un rythme considérablement plus faible ; le cycle de l’azote est cependant unique en ce sens qu’il dépend presque exclusivement de réactions redox régulées par des micro-organismes. Par conséquent, les microbes sont non seulement essentiels au maintien du cycle, mais également à la biodisponibilité de l’azote et constituent donc un facteur de contrôle clé de la production primaire. Il est clair que les micro-organismes présents dans les sédiments marins jouent un rôle important dans l’équilibre du cycle mondial de l’azote 6 , mais malgré les progrès récents dans notre compréhension des transformations de l’azote dans l’environnement marin, notre connaissance des principaux les processus, les flux, les taux de renouvellement, les rendements énergétiques, les répartitions spatiales et la taille des populations dans les sédiments des grands fonds sont étonnamment limités. L’une des principales raisons de cette limitation est l’accent historique mis sur les sédiments riches en matières organiques sur les marges continentales ou dans les zones d’upwelling. Dans de tels systèmes, la disponibilité d’énergie, sous forme de carbone organique, s’accompagne d’une activité microbienne élevée, limitant la pénétration de l’oxygène et des nitrates à une profondeur de quelques centimètres ou moins. Ainsi, l’importance de la transformation de l’azote dans les sédiments à forte charge organique est éclipsée par d’autres réactions redox telles que la réduction des sulfates, pour lesquelles les zones de réaction peuvent s’étendre jusqu’à des profondeurs de plusieurs centaines de mètres. Cependant, à mesure que les chercheurs portent leur attention sur les vastes régions oligotrophes du fond océanique, il devient évident que la pénétration profonde de l’oxygène et la persistance des nitrates dans la colonne de sédiments représentent un scénario géochimique répandu qui diffère nettement de la séquence redox diagénétique classique des sédiments riches en matières organiques. sédiments. Considérant que plus de la moitié du fond océanique est recouverte de sédiments oligotrophes, ces nouvelles connaissances suggèrent que la transformation de l’azote est un processus omniprésent dans la biosphère sédimentaire profonde. Le déséquilibre des activités métaboliques des nitrifiants et des dénitrifiants maintient des concentrations élevées de nitrate dans toute la colonne de sédiments et est probablement dû à une disponibilité limitée de carbone pour les dénitrifiants. Le résultat est un efflux de nitrate dans l’eau de mer sus-jacente et dans l’aquifère basaltique sous-jacent, deux habitats où les nitrates peuvent agir comme un élément nutritif limitant.
Résumé schématique de la transformation de l’azote et des changements de la communauté nitrifiante dans les sédiments oligotrophes avec un socle hydrologiquement actif. L’eau de mer oxygénée des fonds océaniques et de l’aquifère crustal provoque un afflux d’oxygène dans la colonne de sédiments par le haut et par le bas. La minéralisation rapide du carbone organique dans les sédiments de surface libère de l’ammoniac, qui est instantanément consommé par les nitrifiants, favorisant ainsi leur abondance et leur diversité initiales élevées. Sous les couches superficielles de haute activité, l’abondance et la diversité des nitrifiants diminuent en raison d’une minéralisation restreinte du carbone organique et donc d’une disponibilité limitée de l’ammoniac. Dans la zone de transition oxique-anoxique (OATZ), l’ammoniac est à nouveau rendu disponible par diffusion à partir des couches anoxiques plus profondes qui soutiennent la minéralisation anaérobie et la libération d’ammoniac, provoquant une augmentation de l’abondance et de la diversité des cellules nitrifiantes dans l’OATZ. Dans la zone anoxique, la nitrification est inhibée et la diversité et l’abondance des nitrifiants diminuent. En entrant dans la zone de transition anoxique-oxique (AOTZ),les nitrifiants ne sont plus limités en oxygène, et on détecte une augmentation en abondance mais pas en diversité. À mesure que les nitrifiants quittent l’AOTZ, leur abondance diminue et leur diversité reste faible. Bien que la dénitrification se produise dans toute la colonne de sédiments, la disponibilité limitée de carbone organique est probablement responsable du maintien des taux de dénitrification à un niveau inférieur à la production de nitrate, ce qui entraîne un efflux de nitrate dans le fond océanique sus-jacent et dans la croûte sous-jacente.
La dénitrification est généralement un processus par étapes de respiration anaérobie chez les hétérotrophes qui peuvent utiliser des oxydes d’azote comme accepteurs d’électrons terminaux grâce à des enzymes codées par une suite de gènes connus. La première étape de la réduction du nitrate (NO 3 – ) en nitrite (NO 2 – ) se produit via la NO 3 – réductase périplasmique codée par nap ou la NO 3 – réductase liée à la membrane codée par nar . La deuxième étape, la réduction du NO 2 en oxyde nitrique (NO), est catalysée par deux NO 2 – réductases fonctionnellement similaires mais structurellement distinctes, une NO 2 – réductase contenant du cuivre et un cytochrome cd 1 contenant du NO 2 – réductase codé par nirK et nirS , respectivement. La réduction du NO en oxyde d’azote (N 2 O) s’effectue via des NO réductases codées par divers gènes nor, et le N 2 O est finalement réduit en gaz N 2 inerte par des N 2 O réductases codées par nos. NO peut également être disproportionné en N 2 et O 2 par le gène putatif de l’oxyde nitrique dismutase codé par nod dans un processus de dénitrification oxygénée. Il a été démontré que la dénitrification est un processus modulaire, certains micro-organismes étant capables d’effectuer la réduction complète du NO 3 – en N 2 tandis que d’autres ne sont capables que d’une ou d’un sous-ensemble d’étapes. De plus en plus, les efforts de séquençage démontrent que divers dénitrifiants partiels constituent la majorité des dénitrifiants environnementaux, mais la dénitrification est souvent traitée comme un processus complet et singulier en biogéochimie marine. https://www.nature.com/articles/s43705-023-00284-y
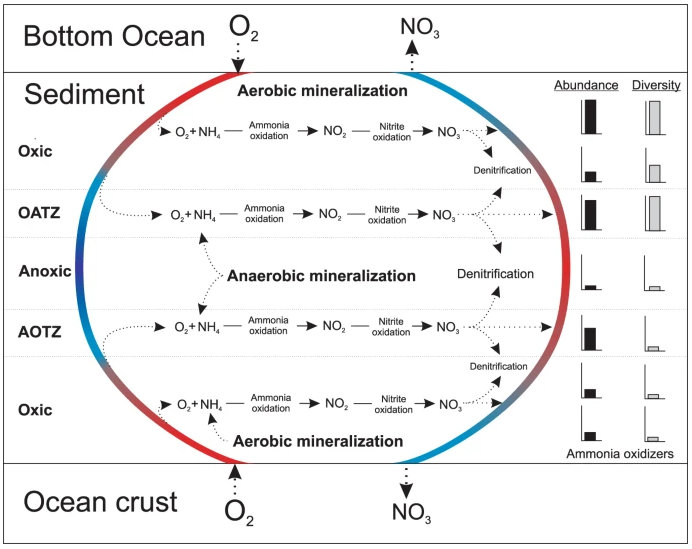
L’océan abrite une communauté microbienne diversifiée et métaboliquement polyvalente qui effectue les transformations biochimiques complexes qui déterminent le cycle de l’azote, notamment les processus de fixation, d’assimilation, de nitrification et de perte d’azote. Le réchauffement des océans, l’acidification, l’enrichissement en nutriments et la stratification de l’eau de mer ont des effets majeurs sur le cycle microbien de l’azote, mais la désoxygénation généralisée des océans est peut-être la conséquence la plus importante pour les micro-organismes impliqués dans les voies de transformation de l’azote aérobie et anaérobie. https://www.nature.com/articles/s41579-022-00687-z
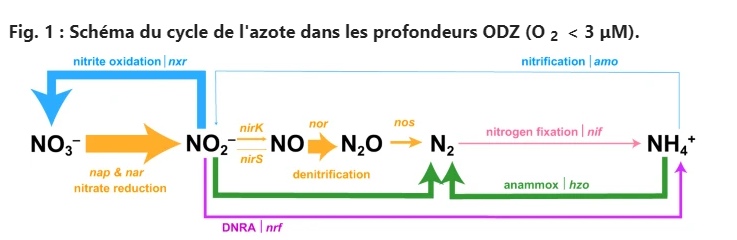
Notre compréhension de la façon dont cet élément est cyclé sur Terre a radicalement changé au cours des dernières années. quelques années. Ici, N existe sous de multiples états d’oxydation et formes chimiques, et est rapidement converti par les micro-organismes terrestres et marins. Jusqu’à récemment, le cycle global de l’azote était considéré comme essentiellement « linéaire » depuis l’atmosphère et vice-versa. Le plus grand réservoir de N sur Terre est le gaz N 2 à triple liaison (78 % de l’atmosphère) et doit être fixé par des micro-organismes avant d’être facilement utilisable par d’autres organismes. N existe dans son état le plus réduit au sein des organismes, mais est rapidement nitrifié en nitrate (par voie aérobie) lorsqu’il est libéré après la mort cellulaire et la lyse. Le nitrate est à son tour dénitrifié en gaz N 2 dans des conditions suboxiques à anoxiques, complétant ainsi le cycle. Depuis le milieu des années 1960, les océanographes ont reconnu des déficits généralisés en ammonium dans les bassins anoxiques, ce qui laissait entendre que l’ammonium pourrait être éliminé par l’activité microbienne anaérobie. La première preuve de l’oxydation anaérobie de l’ammonium (anammox) en gaz N 2 a été obtenue à partir de bioréacteurs anoxiques (dénitrifiants) d’usines de traitement des eaux usées (STEP). De nouveaux organismes liés aux Planctomycetales étaient capable d’oxyder l’ammonium en utilisant du nitrite (plutôt que de l’O 2 ) comme accepteur d’électrons. Il a été conclu que l’oxydation anaérobie de l’ammonium est un nouveau processus dans lequel l’ammonium est oxydé avec du nitrate servant d’accepteur d’électrons dans des conditions anaérobies, produisant du diazote gazeux. Ce procédé biologique a reçu le nom d’« Anammox » (oxydation anaérobie de l’ammonium) et a été breveté. Convenant à des micro-organismes capables d’un métabolisme aussi nouveau, ces bactéries « anammox » possèdent un certain nombre de caractéristiques véritablement uniques, notamment l’utilisation de l’hydrazine (N 2 H 4 , c’est-à-dire le carburant de fusée) comme intermédiaire catabolique libre. Les quatre genres actuellement reconnus de bactéries anammox – Candidatus « Brocadia », « Kuenenia », « Scalindula » et « Anammoxoglobus » – partagent ces caractéristiques physiologiques et morphologiques uniques. Sans reminéralisation du NH 4 + de la matière organique via la dénitrification, on ne sait pas comment l’anammox pourrait être maintenu ; une possibilité est que les hétérotrophes microaérobies soient responsables de la régénération de l’azote dans des conditions d’oxygène faibles. Étonnamment, les concentrations de crénarchéol étaient les plus élevées dans la zone sulfurée de la mer, il semble clair que les Crenarchaeota marinsont une flexibilité métabolique et écologique considérable, basée sur leur présence dans des conditions sulfurées et leur succès évident dans des environnements suboxiques. l’interaction entre le cycle de l’azote et le cycle du carbone devient particulièrement marquante. Si la perte d’azote est principalement due au processus autotrophe de l’anammox plutôt qu’à la dénitrification hétérotrophique, qu’arrive-t-il au carbone organique s’il n’est pas reminéralisé via la dénitrification ? https://www.nature.com/articles/ismej20078
On pense que l’élimination des nitrates et des nitrites dépendante du méthane dans les environnements anoxiques repose sur la syntrophie entre les archées ANME-2d et les bactéries du genre « Candidatus Méthylomirabilis », capable de coupler indépendamment l’oxydation du méthane à la réduction des nitrates via le nitrite en diazote. https://www.nature.com/articles/s41564-023-01578-6
Les changements climatiques, physiques et biogéochimiques entraînent une perte et une récupération rapides de l’oxygène dans un écosystème marin.
Les concentrations d’oxygène dissous (OD) façonnent la biogéochimie et la structure écologique des écosystèmes aquatiques ; par conséquent, comprendre comment et pourquoi l’OD varie dans l’espace et dans le temps est d’une importance fondamentale. Nous montrons que l’OD a diminué dans tout le lac marin et s’est ensuite rétabli dans la colonne d’eau supérieure. Ces changements s’accompagnaient de variations de la température de l’eau et étaient corrélés aux changements de vent, de précipitations et surtout de hauteur de la surface de la mer survenus lors de l’événement El Niño-Oscillation australe de 2015-2016. Nos données indiquent que les propriétés et processus climatiques, physiques et biogéochimiques régulaient collectivement l’OD, produisant des rétroactions liées qui ont entraîné le déclin et la récupération de l’OD. Les facteurs à l’origine de la variabilité de l’oxygène ne sont pas bien compris : les variations de la production via la photosynthèse, la consommation respiratoire d’un large éventail d’organismes et de multiples processus et propriétés physiques. peuvent jouer un rôle important dans la variation. degrés. De nombreux écosystèmes aquatiques sont dépourvus d’oxygène dissous (OD) en profondeur en raison d’une combinaison de l’isolement des eaux souterraines de l’atmosphère et de la respiration communautaire (CR) de la matière organique dans l’eau hors de contact avec l’atmosphère. Les changements environnementaux naturels et anthropiques peuvent également affecter la biologie et la physique du cycle et de la dynamique de l’oxygène. Ainsi, à l’échelle du lac dans son ensemble, les changements dans l’OD intégré sont pour la plupart fortement liés à l’activité biologique plutôt qu’aux flux d’oxygène air-eau. Malgré des statistiques de régression relativement faibles, le vent était le principal facteur physique jouant un rôle dans la prévision des changements dans d’autres variables physiques, chimiques et biologiques. https://www.nature.com/articles/s41598-019-52430-z
La diminution de l’oxygène dissous dans les océans pourrait être préjudiciable à la vie marine et aux cycles biogéochimiques. Cependant, prédire la disponibilité future de l’oxygène avec des modèles qui se concentrent principalement sur les valeurs moyennes temporelles et spatiales à grande échelle pourrait conduire à des prédictions incorrectes. Les écosystèmes marins sont fortement influencés par de courtes fluctuations d’oxygène à petite échelle temporelle et spatiale. La modélisation à grande échelle néglige les fluctuations, qui incluent l’apparition généralisée d’une sursaturation élevée en oxygène à l’échelle quotidienne dans les écosystèmes productifs tels que les récifs coralliens, les herbiers marins et les forêts de mangroves, ainsi que l’hétérogénéité spatiale de la disponibilité en oxygène aux échelles microclimatiques. Dans ces microenvironnements temporels et spatiaux, les fluctuations de l’oxygène contrôlent les cycles biogéochimiques et modifient les réponses des communautés, par exemple au stress thermique et à l’hypoxie. Des projections robustes sur l’impact de la désoxygénation prévue des océans et des côtes nécessitent une meilleure compréhension de la dynamique de l’oxygène dissous couplée à des projections réduites des fluctuations de l’oxygène à de petites échelles pertinentes pour les processus et les communautés biogéochimiques marins. Dans l’ensemble, l’étude de la véritable dynamique de l’oxygène dans les habitats marins productifs peut fournir des informations cruciales sur les mécanismes de rétroaction entre le changement climatique et les écosystèmes marins et peut aider à développer des stratégies efficaces de gestion et de conservation. https://www.nature.com/articles/s41561-023-01217-z
La concentration d’oxygène dissous dans les systèmes aquatiques contribue à réguler la biodiversité, la biogéochimie des nutriments, les émissions de gaz à effet de serre et la qualité de l’eau potable. Les baisses à long terme des concentrations d’oxygène dissous dans les eaux côtières et océaniques ont été liées au réchauffement climatique et à l’activité humaine, mais on sait peu de choses sur les changements dans les concentrations d’oxygène dissous dans les lacs. Bien que la solubilité de l’oxygène dissous diminue avec l’augmentation de la température de l’eau, les trajectoires à long terme des lacs sont difficiles à prévoir. Nous constatons qu’une baisse de l’oxygène dissous est généralisée dans les habitats de surface et d’eau profonde. Le déclin des eaux de surface est principalement associé à une solubilité réduite sous des températures d’eau plus chaudes. Nos résultats suggèrent que le changement climatique et la diminution de la clarté de l’eau ont modifié l’environnement physique et chimique. Les diminutions de l’oxygène dissous dans l’eau douce sont 2,75 à 9,3 fois plus importantes que celles observées dans les océans du monde et pourraient menacer les services écosystémiques essentiels des lacs. https://www.nature.com/articles/s41586-021-03550-y
Les modèles océaniques prédisent une diminution de l’inventaire d’oxygène dissous de l’océan mondial de un à sept pour cent d’ici 2100, causée par la combinaison d’une diminution de la solubilité de l’oxygène induite par le réchauffement et d’une ventilation réduite des profondeurs océaniques. On pense qu’une telle baisse de la teneur en oxygène des océans pourrait affecter les cycles des nutriments océaniques et l’habitat marin, avec des conséquences potentiellement néfastes pour la pêche et les économies côtières. Les données d’observation régionales indiquent une diminution continue des concentrations d’oxygène dissous océanique dans la plupart des régions de l’océan mondial. Cette étude révèle que la teneur mondiale en oxygène des océans a diminué de plus de 2 % au cours des cinq dernières décennies, mais qu’il existe de grandes variations dans les pertes d’oxygène dans les différents bassins océaniques et en fonction de la profondeur de l’océan. https://www.nature.com/articles/nature21399
Les proliférations de phytoplancton en eau douce affectent la santé publique et les services écosystémiques à l’échelle mondiale. Les effets néfastes de telles proliférations se produisent lorsque l’intensité de la prolifération est trop élevée ou lorsque des espèces de phytoplancton produisant des toxines sont présentes. Les proliférations d’eau douce entraînent des pertes économiques de plus de 4 milliards de dollars par an rien qu’aux États-Unis, principalement en raison des dommages causés à la production alimentaire aquatique, aux loisirs et au tourisme, ainsi qu’à l’approvisionnement en eau potable. Nous constatons que l’intensité maximale de la prolifération estivale a augmenté dans la plupart (68 %) des lacs étudiés, révélant une exacerbation globale des conditions de prolifération. La raison derrière l’augmentation de l’intensité de la prolifération de phytoplancton reste cependant floue, car les tendances temporelles ne suivent pas de manière cohérente les tendances de la température, des précipitations, de l’utilisation d’engrais ou d’autres facteurs précédemment hypothéqués. Nos résultats soutiennent les appels à des efforts de gestion de la qualité de l’eau pour mieux tenir compte des interactions entre le changement climatique et les conditions hydrologiques locales. https://www.nature.com/articles/s41586-019-1648-7
Des études récentes suggèrent que l’aluminium (Al) joue probablement un rôle dans le cycle du carbone océanique en modifiant la fixation biologique du carbone et la décomposition du carbone des diatomées marines. Cependant, il reste spéculatif quant à savoir si l’Al a des effets similaires sur d’autres groupes de phytoplancton écologiquement importants, tels que la cyanobactérie fixatrice d’azote d’importance mondiale, Trichodesmium . le taux de décomposition du Trichodesmium POC (carbone organique particulaire/dissout) et du DOC a ralenti respectivement de 20 à 27 % et de 31 à 62 % au cours de la deuxième phase de désintégration, lorsque le DOC et le POC bactérien ont diminué. L’étude suggère que la fertilisation par l’Al dans l’océan de surface via le dépôt de poussière peut augmenter la fixation nette du carbone et la fixation de l’azote associée par Trichodesmium , et donc l’apport de nouvel azote à la zone euphotique, tandis que l’Al provenant des sédiments peut diminuer le taux de décomposition du Trichodesmium en décomposition. installés au fond de l’océan. https://link.springer.com/article/10.1007/s10533-023-01081-4
Le sulfure de diméthyle (DMS) est une source importante d’aérosols de sulfate marin et joue un rôle important dans la modification des propriétés des nuages. Des simulations climatiques entièrement couplées utilisant des calculs dynamiques d’écosystème marin et de DMS sont réalisées pour estimer les flux de DMS dans divers scénarios climatiques et pour examiner pour la première fois le signe et la force des rétroactions phytoplancton-DMS-climat. Les résultats des simulations montrent de légères différences dans la production et les émissions de DMS entre les scénarios climatiques préindustriels et actuels, à l’exception de certaines zones de l’océan Austral. Il y a des changements évidents dans les concentrations de DMS à la surface des océans à l’avenir, et ils sont attribuables à des changements dans la production et la compétition de phytoplancton entraînés par des mécanismes complexes variant dans l’espace. Les comparaisons entre simulations parallèles avec et sans flux de DMS dans l’atmosphère montrent des différences significatives dans les écosystèmes marins et les domaines physiques. Sans DMS, les effets indirects des aérosols sur les nuages et le forçage radiatif entraînent une diminution du nombre de nuages, une augmentation du rayonnement solaire et un climat beaucoup plus chaud. Phaeocystis , un producteur d’organosulfure particulièrement efficace avec un avantage de croissance dans des conditions climatiques plus froides, peut bénéficier de la production du composé grâce aux effets de refroidissement du DMS dans le système climatique. Nos résultats montrent un couplage étroit entre les cycles du soufre et du carbone. L’absorption du carbone océanique diminue sans émissions de DMS dans l’atmosphère. L’analyse indique une faible rétroaction positive phytoplancton-DMS-climat à l’échelle mondiale, avec de grandes variations spatiales entraînées par des groupes fonctionnels autotrophes individuels et des mécanismes complexes. Le signe et la force de la rétroaction varient selon les états climatiques et les groupes de phytoplancton. Cela souligne l’importance d’un module dynamique sur l’écosystème marin et du mécanisme du cycle du soufre dans les projections climatiques. https://link.springer.com/article/10.1007/s10533-018-0430-5
Les populations insulaires aux ressources limitées ont prospéré à Hawaï pendant plus d’un millénaire, mais sont désormais confrontées à de nouveaux défis agressifs concernant les ressources fondamentales, notamment la sécurité et la durabilité des ressources en eau. Caractériser la communauté microbienne dans les écosystèmes d’eaux souterraines constitue une approche puissante pour déduire les changements dus aux impacts humains dus à la gestion des terres dans les aquifères complexes hydrogéologiques. Dans cette étude, nous étudions comment la géologie et la gestion des terres influencent la géochimie, la diversité microbienne et les fonctions métaboliques. Les cycleurs N et S étaient dominés par un S-oxydant putatif couplé à une dénitrification complète ( Acinetobacter ), significativement enrichi jusqu’à 4 fois comparativement parmi les échantillons regroupés par géochimie. La présence significative d’ Acinetobacter déduit le potentiel de bioremédiation des eaux souterraines volcaniques pour la S-oxydation et la dénitrification couplées sous l’action microbienne, fournissant un service écosystémique aux populations insulaires dépendantes des aquifères souterrains. Les microbes jouent un rôle régulateur dans les interactions alambiquées entre l’hydrologie, la géologie et l’utilisation des terres, ce qui entraîne une biogéochimie des eaux souterraines très variable. Les microbes des aquifères souterrains représentent environ 40 % de la vie microbienne de la Terre et sont cruciaux pour les services écosystémiques, notamment la fourniture d’eau potable. Les communautés microbiennes souterraines améliorent la qualité de l’eau en décomposant les matières organiques et en cyclant l’azote (N), le soufre (S) et le fer (Fe). Contrairement à la structuration redox des systèmes sédimentaires, des réactions redox opposées se produisent simultanément dans des microenvironnements uniques (par exemple, des biofilms, des particules), ce qui entraîne un microbiome d’eau souterraine diversifié dans de grands habitats aquatiques souterrains. En conséquence, l’utilisation de composés inorganiques comme sources d’énergie (chimiolithoautotrophie) est une voie métabolique très courante dans les écosystèmes d’eau souterraine, souvent liée aux cycles du carbone (C), de l’hydrogène (H) et du soufre (S) avec l’oxygène (O) et le nitrate. (NO 3 − ) comme accepteurs d’électrons les plus largement utilisés. Dans l’une des rares études sur le microbiome menée dans les aquifères profonds des eaux souterraines hawaïennes, l’analyse a révélé une grande diversité fonctionnelle caractérisée principalement par des métabolismes chimiolithotrophes. Caractériser la communauté microbienne des écosystèmes d’eaux souterraines est une approche puissante pour déduire les changements géochimiques résultant des impacts humains et de l’utilisation des terres dans les aquifères hydrogéologiquement complexes. Les changements dans la structure et la diversité de la communauté microbienne souterraine peuvent alerter les utilisateurs d’une diminution de la qualité de l’eau, car les microbes présents dans le sous-sol réagissent rapidement aux modifications de leur environnement dues aux changements d’utilisation des terres et/ou aux contaminants, entraînant une diminution de la diversité et des voies biogéochimiques raccourcies. La pollution diffuse provenant des systèmes d’évacuation des eaux usées personnelles sur place (OSDS, puisards) et de l’agriculture sont les deux principales sources de nutriments d’origine humaine qui ont un impact sur les écosystèmes côtiers et ont été reconnues comme des problèmes environnementaux majeurs. L’azote est l’un des contaminants de source diffuse les plus courants dans les eaux souterraines. Les apports de N sont des déterminants importants de la géochimie des eaux souterraines à ions majeurs, et les communautés microbiennes souterraines sont responsables de la régulation des effets du N anthropique et de sa transition vers les eaux de surface ou les écosystèmes côtiers. La dénitrification autotrophe est le processus dominant d’élimination de l’azote dans les eaux souterraines oligotrophes où les composés inorganiques (Fe, S, manganèse, Mn ; hydrogène gazeux, H 2 ) sont oxydés en réduisant le nitrite (NO 2 − ) ou le nitrate (NO 3 − ) en azote gazeux. (N 2 ). De plus, la dénitrification microbienne peut se produire à la fois de manière anaérobie, ce qui était considéré comme le seul processus jusque dans les années 1980, et de manière aérobie, décrite pour la première fois dans une usine de traitement des eaux usées oxydant les sulfures. Les apports humains de N dans les eaux souterraines et leurs effets néfastes sur les écosystèmes côtiers. En plus de N, la présence de sulfate détermine la structure de la communauté dans les systèmes d’eau souterraine et indique souvent un lien entre les substrats et la disponibilité des donneurs d’électrons. La présence d’espèces S telles que le sulfate (SO 4 2− ), les sulfures (S 2− ), les polysulfures (S x O 6 2− ), le thiosulfate (S 2 O 3 2− ) et le sulfite (SO 3 2− ) sont révélateurs des apports de l’activité géothermique ou de l’eau de mer dans les systèmes d’eaux souterraines. La présence de SO 4 2− dans les eaux souterraines est souvent attribuée aux produits de réactions abiotiques, cependant, l’oxydation biologique des sulfures anaérobies et aérobies est thermodynamiquement favorable dans la plupart des environnements. De plus, la disponibilité relative de S, C et Fe réduits sont également des déterminants clés des voies d’élimination des nitrates. Lorsque S est présent dans des systèmes limités en carbone, la dénitrification est alimentée par l’oxydation microbienne du soufre réduit en sulfate et se produit beaucoup plus rapidement que la production abiotique de sulfate. Cependant, il a été démontré que les taux d’oxydation biotique du H 2 S dépassent de loin l’oxydation abiotique dans des conditions aérobies et anaérobies, suggérant en outre que la production de SO 4 2− dans ces échantillons peut être produite par une activité microbienne. Les concentrations de SO 4 2− sont des facteurs importants de la structure de la communauté microbienne. Le genre Acinetobacter a été caractérisé dans les eaux souterraines oligotrophes avec un génome polyvalent et un grand potentiel pour le traitement de l’eau, comme un réducteur d’azote, mais des travaux plus récents ont montré que les membres ont des capacités de nitrification (oxydation de l’ammoniac en nitrate) à l’aide de substrats organiques (hétérotrophes) et de dénitrification aérobie. Plusieurs espèces d’ Acinetobacter ont été décrites dans plusieurs systèmes industriels, notamment le traitement des eaux usées du soufre, du Mn et du H 2. Les souches d’Acinetobacter ont été identifiées pour la première fois dans le processus d’élimination des sulfures dénitrifiants à l’aide de communautés microbiennes capables à la fois d’hétérotrophie et d’autotrophie pour gagner de l’énergie de la S-oxydation avec un ensemble complet de gènes pour réduire le NO 3 2− en N 2. Les électrons nécessaires à la réduction de N proviennent de l’oxydation microbienne du soufre produisant du SO 4 2− par Acinetobacter et éventuellement Thiobacillus. Les impacts humains de l’agriculture et des eaux usées sont des déterminants importants de la géochimie, de la qualité de l’eau et de la structure de la communauté microbienne (en particulier les groupes fonctionnels du cycle N) dans les écosystèmes d’eaux souterraines. Dans cette étude, la fonction microbienne putative d’élimination de l’azote hautement enrichie est la dénitrification complète (nitrification hétérotrophe couplée à une dénitrification aérobie) via l’oxydation du soufre par Acinetobacter. On pense que la disponibilité de carbone organique labile limite la viabilité de l’élimination hétérotrophe de l’azote dans les eaux souterraines. Les effluents d’eaux usées bien oxydés ont également tendance à contenir de plus petites quantités de carbone labile limitant les processus microbiens de réduction de N dans certains environnements. Les découvertes sur le potentiel enrichi de l’oxydation du soufre et de la dénitrification chimolithoautrophiques dans le bassin versant de Hualālai ont des implications pour les gestionnaires de l’eau. Bien que la dénitrification par oxydation des sulfures entraîne une diminution de la charge en N, le métabolisme est potentiellement préjudiciable au fonctionnement des puits ou des pompes. Les concentrations de sulfate, la dureté de l’eau et la corrosion peuvent augmenter, provoquant ainsi la précipitation d’ions hors de la solution lorsque de l’oxygène est rencontré lorsque les eaux souterraines sont pompées vers la surface. Des communications personnelles avec des propriétaires de puits privés le long de la zone du rift ont confirmé que certains puits connaissent des eaux et des précipitations rances et nauséabondes. Cependant, la présence significative d’ Acinetobacter déduit le potentiel de biorestauration des communautés microbiennes des eaux souterraines volcaniques pour éliminer simultanément le N et le S, fournissant ainsi un service écosystémique aux aquifères d’eaux souterraines aux ressources limitées. https://www.nature.com/articles/s43705-023-00261-5
Les bactéries apportent une immense contribution aux processus biogéochimiques dans l’écosystème des eaux souterraines comme dans tout autre écosystème. Par conséquent, une bonne compréhension de la diversité bactérienne est cruciale pour évaluer l’efficacité des politiques de gestion des eaux souterraines. Dans cette étude, nous avons étudié la structure de la communauté bactérienne dans les eaux souterraines de Nouvelle-Zélande à l’échelle nationale à l’aide de l’outil de profilage moléculaire du polymorphisme de longueur des fragments de restriction terminaux (T-RFLP) et déterminé les relations entre la diversité bactérienne et la chimie des eaux souterraines, les paramètres géologiques et l’impact humain. Une diversité bactérienne considérable était présente et les structures de la communauté étaient fortement liées à la chimie des eaux souterraines, et en particulier au potentiel redox et à l’impact humain, reflétant leur influence potentielle sur la détermination de la diversité bactérienne. De plus, le temps de séjour moyen des eaux souterraines a également montré des relations avec la structure de la communauté bactérienne. À ce jour, dans la plupart des régions du monde, la surveillance de l’état de l’environnement SOE a presque complètement négligé la composante microbiologique des systèmes d’eau souterraine. Seule la présence d’ E. coli est régulièrement surveillée dans les eaux souterraines néo-zélandaises, car il s’agit d’une espèce indicatrice de contamination fécale. Les micro-organismes sont la principale force motrice de cet écosystème. Il est donc crucial de comprendre l’état de référence des composants microbiens dans les différents systèmes aquifères, ce qui nous permettra de tester des hypothèses liées aux tendances des relations entre les micro-organismes et l’environnement physico-chimique. Les communautés microbiennes sont sélectionnées et régulées par la nature chimique et physique des eaux souterraines et, inversement, les communautés microbiennes des eaux souterraines médient les réactions redox tout en obtenant l’énergie nécessaire à leur survie, contrôlant ainsi les concentrations dissoutes d’éléments tels que le fer, le manganèse, l’azote, le soufre… https://academic.oup.com/femsec/article/86/3/490/637353?login=false
Le rejet des sources hydrothermales sur le fond marin fournit des sources d’énergie pour des écosystèmes dynamiques et productifs, soutenus par des populations microbiennes chimiosynthétiques. Ces populations utilisent l’énergie gagnée en oxydant les produits chimiques réduits contenus dans les fluides de ventilation pour fixer le carbone et soutenir plusieurs niveaux trophiques. Les décharges hydrothermales sont éphémères et la composition chimique de ces fluides varie dans l’espace et dans le temps, ce qui peut donner lieu à des communautés microbiennes géographiquement distinctes. Dans ces emplacements de ventilation, la concentration de fer dissous était le principal facteur de structure de la communauté. Ces résultats donnent un aperçu de la structure naissante des communautés microbiennes et mettent en lumière le développement de diverses communautés lithotrophiques au niveau des sources hydrothermales. Ces bactéries forment des biofilms microbiens, également appelés tapis microbiens. De nombreux organismes dominants dans les sources hydrothermales riches en hydrogène et en soufre sont regroupés au sein du phylum Campylobacterota et de la classe Campylobacteria, Sulfurimonas, Sulfurovum, Nitritruptor, Thioreductor et Arcobacter étant les genres les plus courants. En revanche, dans les systèmes de ventilation hydrothermales présentant des concentrations élevées de fer réduit, la classe des zétaprotéobactéries est le membre dominant de la communauté. Sulfurimonas spp. sont capables de croître avec des composés de soufre réduit et de l’hydrogène comme donneur d’électrons et en aérobie avec de l’oxygène ou en anaérobie avec du nitrate ou du nitrite comme accepteurs d’électrons. Dans les sites de ventilation dominés par le fer, les zétaprotéobactéries sont considérées comme des ingénieurs de l’écosystème microbien. Cela signifie qu’ils ont le potentiel de moduler le flux de carbone organique vers d’autres microbes et peuvent façonner leur habitat en produisant des minéraux d’oxyhydroxyde de fer et des exopolysaccharides, augmentant ainsi la richesse et la diversité du paysage microbien, affectant la santé et la stabilité de l’environnement, considérées comme des taxons clés, car elles sont nécessaires à la survie d’autres microbes et sont des moteurs de la biodiversité des écosystèmes. Les fluides d’évent axial sont généralement dominés par les sulfures avec des rapports molaires H 2 S/Fe de 2 à > 1 000. En revanche, les fluides d’évent 2 sont dominés par le fer, avec des rapports molaires H 2 S/Fe de 0,01 à < 0,0001. Par conséquent, une analyse réalisée sur les 10 principaux taxons par classe et paramètres chimiques. L’abondance des zétaprotéobactéries était corrélée à la concentration en fer dissous. H 2 S est corrélé aux Campylobactéries et inversement aux Zétaprotéobactéries. Le Mn avait des impacts inverses sur la structure de la communauté que Fe et le pH, tandis que H 2 S était inversement corrélé à la température. Cela montre que la température et l’emplacement des évents ont une influence sur la composition de la communauté microbienne. De plus, les concentrations de fer et de sulfure ont un impact sur la composition de la communauté. il était surprenant que le sulfure ait été déterminé comme étant un facteur plus mineur de la structure de la communauté. À l’inverse, il n’était pas surprenant que le fer dissous soit un moteur de la structure de la communauté. Chaque site présentait une composition distincte de la communauté microbienne, cela confirme l’influence de la biogéographie ainsi que l’impact de la géochimie sur la diversité microbienne. https://academic.oup.com/femsec/article/100/2/fiae001/7515283?searchresult=1
https://journals.asm.org/doi/full/10.1128/aem.01374-15
https://ami-journals.onlinelibrary.wiley.com/doi/full/10.1111/1462-2920.14011
Des milliers de génomes microbiens mettent en lumière les processus biogéochimiques interconnectés dans un système aquifère
https://www.nature.com/articles/ncomms13219
Le monde souterrain héberge jusqu’à un cinquième de toute la biomasse, y compris les communautés microbiennes qui sont à l’origine des transformations essentielles aux cycles biogéochimiques de la Terre. Cependant, on sait peu de choses sur la structure des communautés microbiennes complexes dans de tels environnements et sur la manière dont les interactions inter-organismes façonnent le fonctionnement des écosystèmes. Les micro-organismes conduisent les transformations des composés organiques et inorganiques dans cet environnement et contrôlent ainsi les cycles biogéochimiques. On sait peu de choses sur la façon dont les organismes des écosystèmes souterrains sont interconnectés métaboliquement. les organismes sont liés via des transferts métaboliques : le transfert de produits de réaction redox d’un organisme à un autre. Cependant, aucun environnement complexe n’a été suffisamment disséqué pour résoudre les réseaux d’interactions métaboliques qui les sous-tendent. Cela restreint la capacité des modèles biogéochimiques à capturer des aspects clés des cycles du carbone et d’autres nutriments.
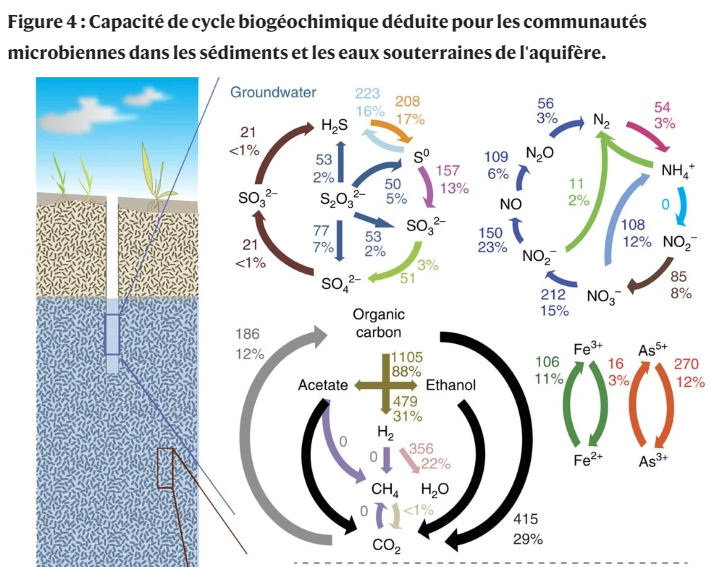
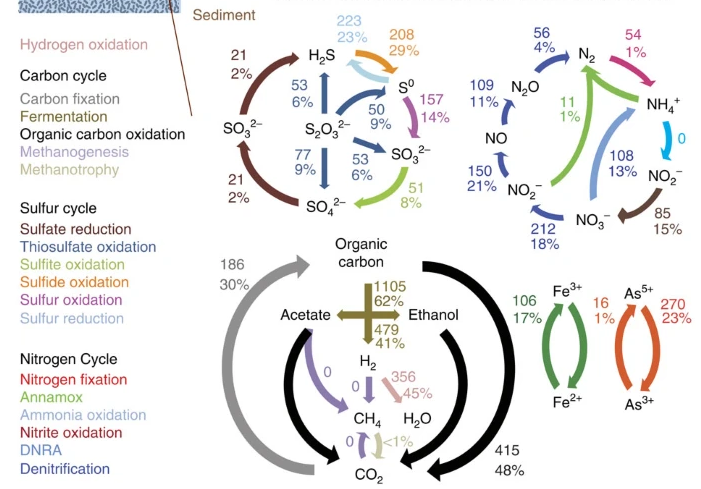
Le sulfure est largement distribué dans le milieu aquatique, mais a été largement négligé en tant que facteur environnemental pour les organismes aquatiques. Le sulfure à des concentrations nanométriques à millimolaires affecte négativement la cytochrome c oxydase, diverses autres enzymes, les protéines de transport de l’oxygène, les structures cellulaires et, par conséquent, les fonctions physiologiques des organismes. https://www.sciencedirect.com/science/article/abs/pii/0166445X9290015F?via%3Dihub
Disulfure de carbone (CS2), le sulfure de carbonyle (OCS), le sulfure de diméthyle (DMS) et le sulfure d’hydrogène (H2S) sont des traces de gaz soufrés trouvées dans la plupart des échantillons d’eau naturelle atmosphérique et de surface. Ils sont tous impliqués dans la formation de dioxyde de soufre (SO2) dans la stratosphère et/ou la troposphère, et pourrait donc jouer un rôle dans le changement climatique mondial.
Le sulfure en tant que phytotoxine du sol
https://www.frontiersin.org/articles/10.3389/fpls.2013.00268/full
Dans les sols des zones humides et les sédiments sous-marins des systèmes marins, saumâtres et d’eau douce, le sulfure de phytotoxine puissant peut s’accumuler à la suite de la réduction microbienne du sulfate au cours de l’anaérobiose, son niveau dépendant des conditions édaphiques dominantes. Dans cette revue, nous comparons un vaste corpus de littérature sur les effets phytotoxiques de ce composé de soufre réduit dans différents types d’écosystèmes, et passons en revue les effets du sulfure à plusieurs niveaux d’écosystème : le fonctionnement écophysiologique de plantes individuelles, les associations plantes-microbes et les effets sur la communauté. y compris les interactions de compétition et de facilitation. Des publications récentes sur les interactions multi-espèces dans la rhizosphère montrent des mécanismes encore plus complexes expliquant la résistance aux sulfures. Il est conclu que le sulfure est une phytotoxine puissante, affectant profondément la condition physique des plantes et le fonctionnement des écosystèmes dans toute la gamme des types de zones humides, y compris les systèmes côtiers. De plus, les températures plus élevées résultant du changement climatique mondial pourraient entraîner une augmentation des taux de production de sulfures dans les eaux peu profondes. Si le sulfure produit ne peut pas être suffisamment séquestré dans le sol par des métaux tels que le fer, le sulfure libre (dissous) s’accumulera. Les concentrations de sulfures dans les eaux interstitielles des sédiments varient largement jusqu’à 15 mmol L −1 dans les sédiments marins. Le composé de soufre réduit agit comme une phytotoxine puissante (tout aussi toxique que le cyanure), en inhibant l’activité de la cytochrome c oxydase dans les mitochondries, conduisant à un blocage ultérieur de la production d’énergie et en affectant négativement une gamme d’autres enzymes contenant des métaux. Le H 2 S gazeux prédomine normalement sur les deux formes ioniques dans les systèmes d’eau douce, car le pH de la plupart des sols anaérobies est tamponné autour de 6 à 7 en raison du HCO − Mécanisme tampon du CO 2 , entraînant des abondances relatives de H 2 S de 95 à 60 %. Dans les systèmes marins, cependant, le pH est souvent autour de 7,5, ce qui conduit à une abondance relative de seulement 30 % pour H 2 S et 70 % pour HS −. Dans les écosystèmes marins et saumâtres, les concentrations de sulfate sont 10 à 1 000 fois plus élevées que dans les systèmes d’eau douce, stimulant les réducteurs de sulfate qui jouent un rôle imminent dans la décomposition et la production concomitante de sulfures. Ainsi, le rôle du sulfure en tant que toxine naturelle potentielle dans les sédiments salins est bien établi. La toxicité du sulfure peut également survenir lorsque les niveaux et les apports de soufre restent inchangés, mais une charge accrue en matière organique augmente les taux de réduction du sulfate en fournissant des donneurs d’électrons issus de sa décomposition. En raison du forçage anthropique, les plantes des zones humides d’eau douce et des systèmes aquatiques sont aujourd’hui confrontées à des concentrations de soufre beaucoup plus élevées à l’échelle mondiale. De plus, la charge en sulfate des eaux souterraines a augmenté en raison de l’oxydation anaérobie des eaux souterraines. composés soufrés réduits par couplage chimiolithoautotrophique de l’oxydation des sulfures et de la réduction des nitrates dans les bassins versants et les zones humides chargés en nitrates. La toxicité des sulfures sera beaucoup moins fréquente pour les macroalgues que pour les plantes vasculaires, car elles ne possèdent que des rhizoïdes et ne font pas saillie dans les sédiments anoxiques, mais sont souvent attachées à des substrats tels que la roche et le corail. À marée basse, cependant, les sulfures peuvent s’accumuler sous des tapis denses de macroalgues. À titre d’exemple, des conditions anoxiques et des niveaux élevés d’ammonium ont été mesurés dans des tapis de Cladophora , entravant la croissance des herbiers marins. On peut s’attendre à ce qu’il ait également conduit à une accumulation de sulfure, comme le montrent les tapis d’Ulva dans les lacs côtiers. Le sulfure ne peut s’accumuler à des concentrations élevées dans les eaux de surface que si l’eau est anoxique et si son oxydation est empêchée. On sait que le sulfure est capable d’entraver l’absorption des nutriments par les plantes, ce qui n’est pas surprenant compte tenu de sa perturbation fondamentale du métabolisme cellulaire et du transfert d’énergie. De plus, la perte de racines due à la mort et la diminution concomitante du rapport racines/pousses conduisent à une absorption déséquilibrée des nutriments. Le sulfure peut altérer l’absorption de l’azote, phosphore et du fer. Ensuite, la production d’acide résultant de l’oxydation des sulfures peut entraîner une perte de Mg et de Ca des sites d’échange de cations du sol dans la rhizosphère, et une moindre disponibilité concomitante de ces macro-ions. Comme notre aperçu montre que même de faibles concentrations de sulfure sont capables d’interagir avec la biogéochimie des nutriments, il est clair que le sulfure peut être un moteur puissant des processus et du fonctionnement des écosystèmes, également en relation avec l’évolution des équilibres S mondiaux. Les recherches futures devraient inclure les interactions entre les plantes, les communautés microbiennes, la faune et la chimie du sol, afin de bien comprendre et expliquer les différences entre les réponses des plantes, de la végétation et des écosystèmes aux sulfures.
La toxicité du H2S a été attribuée à sa capacité à inhiber la cytochrome c oxydase de la même manière que le HCN. Cependant, l’utilisation réussie de la méthémoglobine pour le traitement des intoxications au HCN n’a pas été efficace pour les intoxications au H2S, même si le groupe hème ferrique de la méthémoglobine récupère le H2S. L’incubation d’hépatocytes isolés avec des solutions de NaHS (une source de H 2 S) a entraîné une déplétion en glutathion (GSH). De plus, une déplétion en GSH a également été observée dans le tampon TRIS-HCl (pH 6,0) traité avec NaHS. Plusieurs chélateurs ferriques (desféroxamime et DETAPAC) et enzymes antioxydantes (superoxyde dismutase [SOD] et catalase) ont empêché la déplétion du GSH acellulaire et hépatocytaire. Les hépatocytes appauvris en GSH étaient très sensibles à la cytotoxicité du NaHS, ce qui indique que le GSH détoxifiait le NaHS ou le H 2 S dans les cellules. La cytotoxicité a également été partiellement évitée par la desféroxamine et le DETAPC, mais elle a été augmentée par l’EDTA ferrique ou l’EDTA. Le H 2 S peut réduire le fer ferrique lié intracellulaire pour former du fer ferreux non lié, qui active le fer. De plus, H 2 S peut augmenter la formation hépatocytaire d’espèces réactives de l’oxygène (ROS) (connues pour se produire avec la chaîne de transport d’électrons). La cytotoxicité du H 2 S implique donc également une espèce soufrée réactive, qui épuise le GSH et active l’oxygène pour former des ROS. https://www.tandfonline.com/doi/full/10.1080/03602530600959607?src=recsys
Un examen complet des traitements contre l’intoxication au sulfure d’hydrogène : passé, présent et futur
https://www.tandfonline.com/doi/full/10.1080/15376516.2022.2121192?src=recsys
L’intoxication au sulfure d’hydrogène (H 2 S) reste une source importante de décès professionnels et constitue la deuxième cause la plus fréquente de décès induits par les gaz toxiques. Il s’agit d’un toxique systémique rapidement métabolisé ciblant les mitochondries, entre autres organites. L’intoxication est généralement aiguë, mais des scénarios d’exposition chroniques ou intermédiaires se produisent également. Certains défauts génétiques du métabolisme du H 2 S conduisent à un empoisonnement chronique mortel au H 2 S. Lors d’expositions aiguës, les systèmes neuronal, respiratoire et cardiovasculaire sont les principaux organes cibles, entraînant une détresse respiratoire, des convulsions, une hypotension et des irrégularités cardiaques. Certains survivants d’une intoxication aiguë développent des séquelles à long terme, notamment au niveau du système nerveux central. Cependant, des anomalies génétiques dans le métabolisme du H 2 S produit de manière endogène en métabolite sulfate non toxique, comme celui provoqué par un déficit en ETHE1, ont été associées à une intoxication chronique au H 2 S. Le déficit en ETHE1 et des anomalies génétiques similaires provoquent une neurodégénérescence chronique induite par le H 2 S, résultant de l’accumulation de H 2 S conduisant à la mort au cours de la première décennie de la vie. La principale lésion biochimique de l’intoxication au H 2 S, largement reconnue, est l’inhibition de l’enzyme cytochrome C oxydase au sein de la chaîne de transport d’électrons dans les mitochondries, bloquant la synthèse de l’ATP. Les lésions biochimiques comme l’inhibition de la cytochrome c oxydase et les lésions pathologiques résultant d’une exposition répétée au H 2 S sont cumulatives. Après absorption, le H 2 S subit une oxydation rapide de phase I dans les mitochondries qui conduit à une détoxification. Il est principalement oxydé par les voies d’oxydation des sulfures dans les mitochondries. Trois enzymes ont été impliquées dans la détoxification du H 2 S, dont le sulfure : la quinone oxydoréductase (SQOR), la sulfure dioxygénase (SDO/ETHE1) et la rhodanese, une sulfurtransférase : SQOR est une enzyme liée à la membrane qui catalyse l’oxydation des sulfures en soufre sulfane et réduit ses disulfures de cystéine produisant des groupes persulfure (SQR-SSH). Après SQOR, SDO/ETHE1 catalyse l’oxydation du groupe sulfane-soufre et persulfure produisant du sulfite (H 2 SO 3 ). La soufre transférase rhodanese et SQOR transfèrent ensuite les ions soufre de SQR à H 2 SO 2 pour synthétiser le thiosulfate (H 2 S 2 O 3 ). La thiosulfate réductase réduit davantage H 2 S 2 O 3 en sulfate (H 2 SO 4 ). Il est important de noter que l’oxydation du H 2 S et la production de thiosulfate nécessitent également 1,5 mole d’O 2 pour chaque mole de H 2 S provenant de la chaîne de transport d’électrons, réduisant encore davantage l’apport d’O 2. Le thiosulfate et le sulfate peuvent tous deux être excrétés par les reins. Lorsque les concentrations de H 2 S dans les mitochondries dépassent la vitesse à laquelle le gaz peut être métabolisé, il devient cytotoxique comme dans le cas du déficit en ETHE1 qui sera discuté plus tard sous les effets chroniques de l’exposition au H 2 S. Le H 2 S subit également un métabolisme de phase II par la thiol-S-méthyltransférase, qui est responsable de la méthylation du H 2 S en méthanethiol (CH 3 SH) et en sulfure de diméthyle (CH 3 SCH 3 ). Le résultat net est l’ajout de deux groupes méthyle et l’élimination des ions hydrogène. L’activité de la thiol-S-méthyltransférase s’est avérée la plus active dans le foie et les intestins, bien que cette voie métabolique soit considérée comme 10 000 fois plus lente que les voies d’oxydation. Ceci est mis en évidence par le fait que les défauts d’oxydation du H 2 S dans les mitochondries conduisent à une intoxication chronique au H 2 S, ce qui suggère que la voie de méthylation ne contribue pas de manière significative au métabolisme et à l’élimination du H 2 S.
Il est bien établi que le H 2 S inhibe l’enzyme cytochrome C oxydase dans la chaîne de transport d’électrons, conduisant à l’inhibition de la production aérobie d’ATP. Cela conduit à une augmentation de la respiration anaérobie. La réduction de la respiration aérobie provoque une acidose lactique, une caractéristique de l’empoisonnement au H 2 S, entraînant des effets cytotoxiques entraînant un stress oxydatif et la mort cellulaire. Les tissus qui dépendent fortement de la phosphorylation oxydative, comme le cerveau et le cœur, sont particulièrement sensibles. L’exposition au H 2 S peut déclencher le processus de nécrose oncotique du fait que les cellules sont incapables de reconstituer les caspases responsables de la mort cellulaire programmée et de la déplétion énergétique suite à l’inhibition de la cytochrome C oxydase. Même si le H 2 S est rapidement métabolisé, une seule exposition aiguë supprime généralement l’activité de la cytochrome C oxydase pendant au moins 48 à 72 h, ce qui suggère qu’un manque d’ATP résultant de l’inhibition de cette enzyme clé contribue à la mort cellulaire et à la pathologie des organes. Le H 2 S cible principalement le système neurologique. Le ou les mécanismes exacts de la toxicité du H 2 S sur le système nerveux central sont probablement complexes et ne sont pas entièrement connus. Les effets immédiats comprennent le stress oxydatif et la dérégulation des neurotransmetteurs. La dopamine, la sérotonine et la noradrénaline augmentent lors de l’exposition au H 2 S. Plusieurs études épidémiologiques ont été menées pour étudier l’exposition chronique au H 2 S, les individus exposés au H 2 S présentaient une incidence plus élevée d’irritation respiratoire, et certains présentaient même une activité diminuée de la cytochrome c oxydase, incidences plus élevées d’asthme, de bronchite chronique, de maladie pulmonaire obstructive chronique et de méthémoglobinémie.
Nous avons mentionné précédemment des défauts génétiques qui conduisent à une altération du métabolisme du H 2 S. Les défauts génétiques ETHE1 provoquent une encéphalopathie éthylmalonique, une maladie neurodégénérative autosomique récessive qui entraîne la mort au cours de la première décennie de la vie. Ce défaut est caractérisé par une élévation du H 2 S dans les muscles et les tissus cérébraux conduisant à une toxicité pour le cerveau en raison d’un défaut du métabolisme du H 2 S dans les mitochondries . L’accumulation de H 2 S est due à l’absence de sulfure dioxygénase, une des enzymes responsables de la détoxification du H 2 S dans les mitochondries . Les déficits en MOCS1, MOCS2 et GPHN sont également associés à un déficit en cofacteur de molybdène (MoCD) . Le MoCD est également un trouble autosomique récessif caractérisé par des déficits en sulfite oxydase, en xanthine oxydoréductase et en aldéhyde oxydase, dont certains conduisent à une altération du métabolisme du H 2 S. Comme le déficit en ETHE1, le métabolisme du H 2 S est affecté par l’activité déficiente de la sulfite oxydase, oxydant le sulfite en sulfate moins toxique . Une augmentation de H 2 S et de sulfites conduit à une inhibition cumulative chronique de l’enzyme cytochrome C oxydase. Non seulement il y a une augmentation du H 2 S et des sulfites, mais une augmentation du lactate et du thiosulfate est également observée dans les structures cérébrales des patients. Les déficits en ETHE1 et MOCS1 entraînent une mort précoce. Les patients atteints d’encéphalopathie éthylmalonique et de MoCD présentent des retards de développement et une dégénérescence des noyaux gris centraux, du tronc cérébral, du cortex cérébral, du cervelet et des corps calleux . Améliorer et prolonger la vie des patients atteints nécessite de combler les lacunes en matière de connaissances dans le traitement de la toxicité chronique sous-jacente du H2S.
Sulfate dans les écosystèmes d’eau douce : examen des sources, des cycles biogéochimiques, des effets écotoxicologiques et de la bioremédiation
https://www.sciencedirect.com/science/article/abs/pii/S001282522030492X
L’augmentation des concentrations de SO 4 2- dans les systèmes d’eau douce influence les processus biogéochimiques du carbone, de l’azote et du phosphore. De même, la disponibilité du fer peut être cruciale pour déterminer les effets néfastes du SO 4 2- sur les récepteurs environnementaux. La littérature examinée démontre clairement que la pollution par le SO 4 2- peut avoir des effets toxiques sur les plantes aquatiques et les organismes animaux, notamment les poissons, les invertébrés et les amphibiens, et qu’elle peut également avoir des implications négatives sur la santé humaine. Les systèmes de bioremédiation offrent des possibilités d’atténuer les impacts du SO 4 2- , mais les efficacités d’élimination varient largement de 0 % à 70 % dans les systèmes de traitement tels que les zones humides artificielles.. Nous concluons qu’il est urgent d’examiner l’augmentation des concentrations et des flux de SO 4 2- à différentes échelles spatiales, car la perturbation mondiale actuelle du cycle du S est susceptible d’être accélérée par le changement climatique et les activités de développement humain. Les effets néfastes de cette situation sur les organismes d’eau douce du monde entier pourraient s’avérer préjudiciables au bien-être futur des humains et des écosystèmes. Malgré des réductions significatives des dépôts atmosphériques de soufre (S) d’origine anthropique dans de grandes parties de l’Amérique du Nord et de l’Europe au cours des dernières décennies, les concentrations actuelles de SO 4 2- restent supérieures aux niveaux de fond naturels dans les eaux de surface et souterraines et ont même augmenté depuis le milieu de l’année 20e siècle. Les impacts de l’augmentation des concentrations de SO 4 2- sont mal compris, notamment en ce qui concerne plusieurs aspects des implications sur la structure et le fonctionnement des écosystèmes d’eau douce, même si les effets néfastes sur les systèmes d’eau douce sont bien connus, tels que l’abaissement du pH en combinaison avec mobilité accrue des métaux lourds ainsi que de la toxicité des nutriments et des sulfures. Cette section résume les effets du SO 4 2- sur le cycle des éléments majeurs dans les écosystèmes d’eau douce. Plus précisément, nous nous concentrons sur la perturbation des processus biogéochimiques impliqués dans le cycle du carbone, de l’azote et du phosphore dans les systèmes d’eau douce, résultant soit d’une augmentation des concentrations de SO 4 2- , soit d’une augmentation des concentrations de H 2 S. D’autres types de perturbations, comme l’augmentation des taux de méthylation du mercure.
De faibles concentrations de sulfures affectent les transformations des nitrates dans les sédiments des bassins de rétention côtiers d’eau douce et salins
https://www.sciencedirect.com/science/article/abs/pii/S0038071709000182
Les zones côtières sont sujettes aux fortes tempêtes qui peuvent provoquer un afflux d’eau salée dans les sédiments aquatiques d’eau douce. Ces changements dans les conditions environnementales peuvent avoir un impact sur les processus sédimentaires, notamment le cycle de l’azote (N). Les capacités relatives des communautés microbiennes sédimentaires de deux étangs de rétention de terrains de golf d’eau douce qui se déversent dans les zones humides adjacentes, et de deux étangs de zones humides salines proximales, à éliminer les nitrates ont été comparées afin d’évaluer si de faibles concentrations de sulfures modifiaient les processus de transformation de l’azote. Les microcosmes ont été incubés avec du NO 3 – seul, et avec du NO 3 – et du H 2 S. L’oxyde nitreux (N 2 O), le nitrite (NO 2 − ), le NO 3 − , l’ammonium (NH 4 + ), le SO 4 2− et les sulfures volatils acides ont été analysés au fil du temps. Bien que les faibles concentrations de H 2 S aient généralement augmenté les taux d’élimination du NO 3- sur tous les sites, les périodes de latence ont été augmentées et la dénitrification a été inhibée par la faible teneur en sulfures dans les sédiments d’eau douce, comme en témoignent les concentrations plus élevées de N 2 O accumulées par rapport à celles des sédiments d’eau douce. les sédiments des zones humides. En plus de l’incapacité des sédiments d’eau douce à convertir le N 2 O en N 2 en l’absence de C 2 H 2 , des concentrations transitoires anormalement élevées de NO 2 -N se sont accumulées dans les échantillons du bassin de rétention. La formation de NH 4 -N a généralement diminué en raison de l’ajout de H 2 S aux sites d’eau douce ; La formation de NH 4 -N a augmenté initialement dans les zones humides, mais elle était plus importante lorsqu’aucun H 2 S n’était ajouté. Les tempêtes qui permettent l’afflux d’eau de mer contenant du SO 4 2− dans les systèmes d’eau douce peuvent modifier les espèces dominantes d’azote produites par la réduction des nitrates . Même de faibles concentrations de sulfure produisaient une dénitrification incomplète et une diminution de la formation de NH4 + dans ces sédiments côtiers d’eau douce. La dénitrification est l’un des processus naturels d’élimination de l’azote par voie microbienne les plus efficaces dans les sédiments côtiers et limite l’accumulation de NO 3 − par sa conversion en azote gazeux (N 2 ) dans la séquence de réduction NO 3 − → nitrite (NO 2 − ) → oxyde nitrique (NO) → oxyde nitreux (N 2 O) → N 2 . Contrairement à la dénitrification, la réduction dissimilatoire du nitrate en ammonium (DNRA) produit de l’ammonium (NH 4 + ) qui retient l’azote dans le système. La disponibilité de matière organique (MO) peut être un facteur limitant pour la DNRA et peut avoir un impact sur les proportions relatives de dénitrification et de DNRA qui se produisent. Les conditions généralement favorables à la DNRA sont une plus grande disponibilité de MO et de carbone labile tel que le glucose. Un potentiel rédox plus faible peut favoriser la DNRA ; de plus, la DNRA était moins inhibée par la présence d’oxygène (0 à 2 %) que par la dénitrification. Le DNRA peut également supplanter la dénitrification à de faibles concentrations de NO 3 − et à des concentrations élevées de sulfures. Le H 2 S, le produit final de la réduction des sulfates, est hautement toxique pour la plupart des micro-organismes et s’est avéré retarder et inhiber les taux de dénitrification potentiels à divers degrés dans les sédiments des environnements d’eau douce et salée. Cependant, le sulfure peut accélérer l’élimination du NO 3 − car le sulfure peut agir comme donneur d’électrons et le NO 3 − comme accepteur d’électrons. Par conséquent, les charges de H 2 S et de N anthropique peuvent avoir un impact sur les proportions relatives de dénitrification et de DNRA, et sur l’élimination ultérieure du NO 3 − .
Différents composés de soufre réduit (H 2 S, FeS, S 2 O 3 2− ) ont été testés comme donneurs d’électrons pour la réduction dissimilaire des nitrates dans des boues de sédiments amendées par des nitrates. Ce n’est que dans les boues enrichies en sulfures libres que le nitrate a été sensiblement réduit en ammoniac (0,30 µmol de NH4+µmol NON3−), avec oxydation concomitante du sulfure en S 0 (0,55 µmol S0µmol S2−). La concentration initiale en sulfure libre apparaît comme un facteur déterminant du type de réduction des nitrates. À des concentrations extrêmement faibles de S 2− libre (sulfures métalliques), le nitrate a été réduit par dénitrification, tandis qu’à des concentrations plus élevées de S 2− , une réduction dissimilatoire du nitrate en ammoniac (DNRA) et une dénitrification incomplète en oxydes d’azote gazeux ont eu lieu. L’inhibition par les sulfures des NO- et N 2 O- réductases est proposée comme étant responsable de la partie motrice du flux d’électrons de S 2− vers NH 4 + . https://www.sciencedirect.com/science/article/abs/pii/0168649696000517
L’effet de l’oxydation des sulfures acides-volatils (AVS) sur le cycle du Fe−S et la qualité de l’eau dans les drains des plaines inondables côtières provenant de paysages de sols acides-sulfatés a été examiné à l’aide de sédiments naturels et de monosulfure de fer synthétique. L’oxydation de l’AVS s’est produite rapidement (demi-temps ≤ 1 h) et a produit du soufre élémentaire (S 8 0 (s) ) et de l’oxyhydroxyde de fer (FeOOH (s) ). Le processus initial d’oxydation rapide de l’AVS s’est produit sans acidification significative ni modification de la composition de la phase aqueuse. Une acidification sévère (pH < 4) ne s’est produite qu’une fois que le S 8 0 (s) a commencé à s’oxyder en SO 4. Nos résultats démontrent, pour la première fois avec des sédiments naturels, une étape d’oxydation AVS significativement tamponnée au pH (presque neutre), le déclencheur de l’acidification étant l’oxydation du S 8 0 (s) . L’acidification a entraîné la libération, en fonction du pH, de grandes quantités d’Al, Mn, Ni et Zn, même si la teneur en métaux des sédiments était faible. https://pubs.acs.org/doi/10.1021/es0520058
La séquestration du soufre dans la pyrite sédimentaire est un puits majeur dans le cycle biogéochimique mondial du soufre et a des impacts conséquents sur les cycles associés du carbone et de l’oxygène. Dans de nombreux environnements sédimentaires , il existe une zone qui libère du H 2 S si elle est traitée avec un acide. Les matériaux qui produisent ce sulfure sont décrits comme des sulfures volatils acides ou AVS et le sulfure qui est dégagé et collecté par ce traitement est appelé sulfure volatil acide (AVS). Au cours des 40 années qui se sont écoulées depuis que RA Berner a défini pour la première fois le concept de sulfure volatil acide, un certain nombre de mythes scientifiques se sont développés concernant la nature et la composition de l’AVS et son rôle dans le cycle mondial du soufre. Bien que l’AVS ait pu remplir son rôle dans les premiers jours exploratoires de l’étude des sulfures sédimentaires, le concept constitue probablement aujourd’hui plus un obstacle qu’une aide pour faire progresser ce domaine. Il est raisonnable de se demander s’il n’est pas temps d’abandonner complètement le concept AVS et de s’attaquer plus directement au comportement des composants les plus importants connus pour appartenir au groupe des espèces sulfurées qui contribuent au sulfure qui se volatilise lorsque de l’acide est ajouté à un sédiment. L’un des aspects déterminants des cycles biogéochimiques mondiaux interdépendants du carbone, de l’oxygène et du soufre est l’élimination du soufre du système océan-atmosphère et son enfouissement dans les sédiments. L’article fondateur du concept d’AVS sédimentaire en tant que composant « défini opérationnellement » des sédiments anoxiques est probablement celui de Berner (1964a). Il a défini l’AVS en termes de source de H 2 S dégagée à partir des sédiments par l’ajout de HCl 1 N. Cette approche mesure à la fois les minéraux sulfurés réactifs au HCl en phase solide et les sulfures dissous dans les eaux interstitielles. l’AVS est une propriété définie « opérationnellement » qui est le H 2 S (AVS-S) qui se dégage lors de la lixiviation acide des sédiments avec du HCl. En tant que tel, le terme AVS en lui-même ne contient aucune stipulation quant aux sources du H 2 S dégagé. Il ne doit donc jamais être utilisé de manière interchangeable, comme c’est la pratique courante, avec FeS sédimentaire car cela implique une source minérale spécifique. Le sulfure de l’AVS est principalement produit par les activités des procaryrotes sulfato-réducteurs (SRP). Les bactéries sulfato-réductrices (Fig. 19) sont bien connues, mais il devient évident que les archées sulfato-réductrices (SRA) ne se limitent pas aux environnements extrêmes. https://www.sciencedirect.com/science/article/abs/pii/S0304420305001039
Lors du remaniement physique ou biologique des sédiments marins superficiels, les oxydes métalliques sont souvent mis en contact avec des sulfures solides et dissous. Des expériences simulant ces processus de mélange démontrent que dans les sédiments naturels, les oxydes de Mn peuvent complètement oxyder les sulfures en phase solide en SO 4 − dans des conditions anoxiques. La principale source de soufre est probablement le sulfure volatil acide. Les minéraux contenant du Mn +4 sont apparemment plus efficaces que le Mn +3 pour stimuler l’oxydation. Il existe peu ou pas de preuves d’une oxydation complète des sulfures par les oxydes de Fe dans des conditions similaires. La réaction est inhibée par le DNP (dinitrophénol) et l’azoture, ce qui implique une médiation biologique par un groupe de bactéries chimiolithotrophes telles que les thiobacilles, possédant un système cytochrome bien organisé, une phosphorylation oxydative couplée à une oxydation des sulfures, et éventuellement une fixation aulolrophique du CO 2 . https://www.sciencedirect.com/science/article/abs/pii/0016703788903353
L’eutrophisation peut stimuler les émissions de composés soufrés volatils (CSV) accompagnées de variations des variables environnementales dans les lacs. les effets de l’eutrophisation sur les émissions de CSV provenant des sédiments lacustres ainsi que les mécanismes sous-jacents restent flous. La matière organique, Fe 3+ , NO 3 − -N et le soufre total ont eu une influence significative sur les communautés microbiennes dans les sédiments. Les lacs eutrophes favorisent la croissance massive d’algues, et la croissance et la décomposition saisonnières des algues peuvent affecter le cycle du soufre. Il a été rapporté que les lacs d’eau douce eutrophes sont une source cruciale d’émission de composés soufrés volatils (CSV), H 2 S, le méthanethiol (CH 3 SH), le disulfure de carbone (CS 2 ), le sulfure de diméthyle ((CH 3 ) 2 S) et le disulfure de diméthyle ((CH 3 )S 2 ). De manière générale, les CSV sont considérées comme la principale source de mauvaises odeurs dans l’eau en raison de leurs faibles seuils olfactifs. Des VSC excessives peuvent causer de graves dommages au système nerveux humain. De plus, les VSC jouent un rôle essentiel dans la formation des nuages, le changement climatique et le cycle mondial du soufre. Par conséquent, comprendre les émissions de CSV provenant des lacs eutrophes est d’une grande importance pour contrôler la pollution des CSV dans les habitats. La croissance et la décomposition des algues sont considérées comme des sources vitales d’émissions de CSV dans les lacs eutrophes. Bien que la croissance et la décomposition des algues puissent favoriser les émissions de CSV, le soufre dérivé des algues se déposerait principalement dans les sédiments, accompagné d’une teneur croissante en sulfate (SO 4 2− ) au cours de la décomposition des algues. Au cours des dernières années, la concentration de SO 4 2− dans les lacs a augmenté de manière significative en raison des activités humaines, notamment dans les lacs eutrophes. Le SO 4 2− et les précurseurs contenant du soufre, comme la méthionine et la cystéine, peuvent être réduits/dégradés en CSV par des micro-organismes métabolisant le soufre, tels que les bactéries réductrices de soufre (SRB), qui peuvent également affecter les émissions de CSV . La réduction des sulfates est l’un des principaux processus responsables des émissions de VSC, dont le SRB est le principal moteur. Les SRB sont généralement des micro-organismes strictement anaérobies, mais ils peuvent tolérer une certaine quantité d’oxygène par agrégation cellulaire. L’activité du SRB augmente avec la teneur en SO 4 2− des sédiments. Le taux de réduction des sulfates diminuait avec la profondeur dans l’ensemble des sédiments, passant de concentrations plus élevées de SO 4 2− près de la surface à des concentrations plus faibles dans la zone profonde. La teneur en SO 4 2− variait avec la profondeur des sédiments, ce qui entraînait des taux de réduction des sulfates différents en raison de diverses abondances et activités de SRB et affectait les émissions de CSV du lac. Le SO 4 2− pourrait favoriser la réduction du Fe 3+ et les émissions de phosphore dans les sédiments lacustres, ce qui aggraverait les niveaux d’eutrophisation et influencerait le métabolisme du soufre. Les bactéries oxydantes du soufre (SOB) peuvent oxyder le sulfure en SO 4 2− ou en intermédiaires tels que le soufre, le thiosulfate et le sulfite pour réduire les émissions de CSV. Les SOB sont affectés par de nombreux facteurs présents dans les sédiments, tels que la profondeur, la matière organique (MO) et les nutriments, et sont généralement détectés dans les couches supérieures et moyennes des sédiments. Dans les sédiments de mangrove, la température, le potentiel redox et le SO 4 2− sont considérés comme des facteurs importants influençant les communautés microbiennes du cycle du soufre, le soufre élémentaire étant le principal moteur du SOB. Les communautés SOB sont sensibles aux changements dans les substances nutritives telles que la MO et l’azote ainsi que les niveaux de salinité et de NH 4 + dans les sédiments. Le taux d’oxydation du soufre le plus élevé est généralement observé dans les sédiments de surface riches en matières organiques. La croissance et la décomposition des algues peuvent non seulement affecter directement les émissions de VSC, mais également avoir une influence indirecte sur les émissions de VSC à travers l’abondance et la communauté de micro-organismes métabolisant le soufre dans les lacs eutrophes. Des sédiments présentant différents niveaux d’eutrophisation, saisons et profondeurs ont été collectés pour détecter le taux de production de VSC, les potentiels d’oxydation du soufre et de réduction des sulfates ainsi que les variables environnementales. https://www.sciencedirect.com/science/article/abs/pii/S004896972302675X
(1) le sulfure en tant que facteur environnemental dans les habitats aquatiques ; (2) le sulfure comme agent toxique ; (3) tolérance aux sulfures des organismes aquatiques ; et (4) des adaptations limitant la toxicité des sulfures. Le sulfure est largement distribué dans le milieu aquatique, mais a été largement négligé en tant que facteur environnemental pour les organismes aquatiques. Le sulfure à des concentrations nanométriques à millimolaires affecte négativement la cytochrome c oxydase, diverses autres enzymes, les protéines de transport de l’oxygène, les structures cellulaires et, par conséquent, les fonctions physiologiques des organismes. Ces effets toxiques sont bien documentés dans la littérature biomédicale et se produisent également chez les organismes aquatiques étudiés. La tolérance aux sulfures varie considérablement parmi les protozoaires, la méiofaune sédimentaire, les polychètes, les bivalves, les crustacés, les poissons marins et d’eau douce et les plantes aquatiques, souvent en corrélation avec les niveaux relatifs de sulfures dans les habitats respectifs. Les organismes aquatiques ont développé diverses adaptations contre la toxicité des sulfures, peut-être plusieurs agissant de concert. La plupart des animaux sont capables d’éviter les sulfures et de s’en échapper, mais ne peuvent pas les exclure du corps. Aucune cytochrome c oxydase résistante aux sulfures n’a été démontrée et la plupart des animaux sont capables d’un certain degré de méabolisme anaérobie. Divers invertébrés sont entrés en associations symbiotiques avec des bactéries oxydant les sulfures. Certains de ces invertébrés immobilisent et transportent le sulfure au moyen de protéines liant les sulfures ou de persulfures dans le sang. La détoxication du sulfure se produit par méthylation, oxydation non spécifique et oxydation enzymatique par les mitochondries. La détoxication oxydative du sulfure en thiosulfate par les mitochondries est commune à plusieurs taxons majeurs (protozoaires, mollusques, téléostéens, mammifères) et est efficace à de faibles concentrations micromolaires de sulfure. Parmi les organismes dépourvus de symbiotes bactériens oxydant les sulfures, les mitochondries peuvent ainsi constituer la principale défense contre les sulfures environnementaux et permettre à l’organisme entier de tolérer des concentrations de sulfures de 2 à 3 ordres de grandeur supérieures à celles qui inhiberaient la cytochrome c oxydase. https://www.sciencedirect.com/science/article/abs/pii/0166445X9290015F
Cueva de la Mora est un lac de mine acide stratifié en permanence et un système modèle pour les études de drainage minier acide extrême (DMA). En utilisant une combinaison de séquençage d’amplicons, de métagénomique et de métatranscriptomique, nous avons effectué une analyse résolue taxonomiquement des contributions microbiennes au cycle du carbone, du soufre, du fer et de l’azote. Nous avons constaté que l’algue verte active Coccomyxa onubensis dominait la couche supérieure et la chimiocline. La chimiocline avait une activité pour l’oxydation du fer (II) réalisée par des populations de Ca . Acidulodesulfobacterium, Ferrovum, Leptospirillium et Armatimonadetes . L’activité prévue pour la réduction du fer (III) n’a été détectée que dans la couche profonde affiliée aux protéobactéries . L’activité de cycle dissimilitoire de l’azote, y compris la fixation de l’azote et la réduction des nitrates, était principalement prévue dans la chimiocline. Les populations archéennes hétérotrophes avec une activité prévue d’oxydation des sulfures liée aux Thermoplasmatales non cultivées dominaient dans la couche profonde. Abondant sulfate-réducteur D esulfomonile et Ca . Les populations d’Acidodesulfobacterium étaient actives dans la chimiocline. Dans la couche profonde, des populations non cultivées des phyla bactériens Actinobacteria, Chloroflexi et Nitrospirae ont contribué à la fois à la réduction des sulfates et à l’oxydation des sulfures. Sur la base de ces informations, nous avons évalué le potentiel de précipitation de minéraux sulfurés dans la couche profonde comme outil d’assainissement. Nous soutenons que la précipitation des sulfures n’est pas limitée par le potentiel génétique microbien mais plutôt par la quantité et la qualité du carbone organique atteignant la couche profonde ainsi que par les ajouts d’oxygène aux eaux souterraines permettant l’oxydation du soufre. L’ajout de carbone organique et de soufre élémentaire devrait stimuler la réduction des sulfates et limiter la réoxydation des minéraux sulfurés. https://www.nature.com/articles/s41396-022-01320-w
Accumulation de N 2 O dans les sédiments estuariens et côtiers : influence du H 2 S sur la réduction dissimilatrice des nitrates
L’oxyde nitreux (N 2 O) est reconnu comme un puissant gaz à effet de serre et un intermédiaire possible dans la destruction de la couche d’ozone stratosphérique. La concentration atmosphérique de N 2 O est actuellement d’environ 310 ppbv et elle augmente à un rythme de 0,6 à 0,9 ppbv an −1. Bien que la concentration atmosphérique de N 2 O soit inférieure à celle du CO 2 et du CH 4 , le N 2 O est considéré comme contribuant au futur changement climatique mondial en raison de son long temps de séjour (130 à 150 ans) et de sa grande capacité d’absorption d’énergie par molécule. C’est pourquoi la production de N 2 O a attiré beaucoup d’attention parmi les chercheurs. La production de N 2 O suit des voies à la fois anthropiques et microbiogènes. Le premier comprend la combustion de biomasse, les activités agricoles et les processus industriels. Le N 2 O est produit par les microbes comme sous-produit de la nitrification, comme intermédiaire lors de la dénitrification. De plus, il existe très peu de données montrant que le N 2 O est produit par la voie de réduction dissimilatoire du nitrate en ammonium (DNRA), mais les mécanismes enzymatiques de la production de N 2 O sont encore incertains. non décrit. La réduction du N 2 O en N 2 est généralement acceptée comme voie d’élimination du N 2 O. L’équilibre entre la production et l’élimination de N 2 O détermine le niveau d’accumulation de N 2 O dans les milieux naturels. Le facteur environnemental régulant l’accumulation de N 2 O n’est cependant pas clair, car la progression simultanée de ces voies microbiennes est compliquée dans les milieux naturels. Pour une estimation précise des émissions de N 2 O dans l’atmosphère, il est crucial d’élucider le mécanisme d’accumulation de N 2 O dans divers environnements.
Nous avons émis l’hypothèse que l’accumulation automnale de N 2 O dans les sédiments de surface était accélérée par la présence de sulfures libres. On a parfois constaté une accumulation significative de N 2 O dans les sédiments marins côtiers plus profonds contenant du H 2 S. De plus, l’ajout de H 2 S a inhibé de manière marquée la réduction du N 2 O dans les suspensions cellulaires ainsi que dans le sol. Nous avons examiné les effets du H 2 S sur la production et l’accumulation de N 2 O par le biais du processus de réduction des nitrates dans les sédiments estuariens et lagunaires des lacs Shinji et Nakaumi. La présente étude a été réalisée en utilisant des suspensions de sédiments et des bactéries productrices de N 2 O isolées des sédiments du lac Shinji. Les effets du H 2 S sur la production et l’accumulation de N 2 O dans les sédiments de surface des lagons côtiers ont été étudiés à l’aide de suspensions de sédiments et de souches isolées de producteurs de N 2 O halophiles. La dénitrification et l’accumulation de N 2 O ont été déterminées respectivement par des incubations anaérobies avec et sans C 2 H 2 . Les activités dénitrifiantes dans les suspensions sédimentaires des deux lacs ont diminué nettement à une concentration de H 2 S de 3 mg S l −1 , tandis que les accumulations de N 2 O dans les suspensions sédimentaires des lacs Shinji et Nakaumi étaient les plus élevées à 75–100 mg S l −1 et 15 à 50 mg S l -1 H 2 S, respectivement. De plus, le H 2 S avait des effets inhibiteurs (environ 30 h) et retardateurs marqués sur la réduction du N 2 O dans les suspensions des deux lacs. Deux souches de producteurs de N 2 O halophiles ont été isolées des sédiments du lac Shinji. Taux d’accumulation de N 2 O par Aeromonas sp. et Vibrio sp. ont été accélérés à 1 à 5 mg S l −1 et 1 à 10 mg S l −1 H 2 S, respectivement. Les modèles de composés azotés inorganiques après les incubations ont révélé que l’accumulation de NH 4 + et NO 2 − , avec N 2 O concomitant, a été accélérée par H 2 S. Ces résultats indiquent que H 2 S joue un rôle clé dans la régulation de N 2 O. accumulation dans les sédiments eutrophes estuariens ou côtiers . En outre, l’accumulation de N 2 O dans les sédiments des lacs Shinji et Nakaumi devrait provenir d’une réduction dissimilatoire des nitrates en ammonium (DNRA) et non seulement d’une dénitrification en présence de H 2 S. Les activités d’accumulation de N 2 O dans les suspensions des lacs Shinji et Nakaumi se sont révélées significativement plus élevées à 75-100 et 15-50 mg S l −1 H 2 S. Ces activités dans les deux lacs étaient environ 4 à 11 fois supérieures à celles dans chaque cas sans H 2 S.
https://www.sciencedirect.com/science/article/abs/pii/S0272771405003914
Cycle actif du soufre dans le sous-sol terrestre profond
https://www.nature.com/articles/s41396-020-0602-x#Sec8
Le sous-sol terrestre profond reste un environnement où la compréhension des métabolismes microbiens existants est limitée. À Olkiluoto, en Finlande, un dépôt géologique en profondeur est en construction pour le stockage final du combustible nucléaire usé. Il est donc essentiel d’évaluer l’impact potentiel du métabolisme microbien, y compris la génération de sulfures, sur la sécurité du dépôt. Nous avons étudié une eau souterraine profonde où le sulfate est présent, mais la géochimie des eaux souterraines suggère une activité microbienne limitée de réduction du sulfate. L’examen de la communauté microbienne au niveau du génome a révélé des micro-organismes dotés de la capacité métabolique de transformations oxydatives et réductrices du soufre. Il a été démontré que les deltaprotéobactéries ont la capacité génétique de réduction des sulfates et éventuellement de dismutation du soufre, tandis que les Rhizobiaceae , les Rhodocyclaceae , les Sideroxydans et les Sulfurimonas oxydent les composés soufrés réduits. Un examen plus approfondi du protéome a confirmé un cycle actif du soufre, servant à la génération et à la croissance d’énergie microbienne. Nos résultats révèlent que ces eaux souterraines pauvres en sulfures abritent une communauté microbienne active de bactéries sulfato-réductrices et oxydantes de sulfures, médiatrices d’un cycle du soufre qui n’a pas été détecté par la seule surveillance géochimique. La capacité des bactéries oxydant les sulfures à limiter l’accumulation de sulfures a été démontrée davantage dans les incubations d’eaux souterraines et met en évidence un puits potentiel pour les sulfures qui pourrait être bénéfique pour la sécurité du dépôt géologique.
Le sous-sol terrestre abrite une proportion importante des cellules microbiennes de la Terre. Dans le sous-sol terrestre, les donneurs d’électrons dérivés de la photosynthèse sont en abondance relativement faible et les gaz géogéniques, H 2 et CH 4 , fournissent des sources d’énergie pour les communautés microbiennes. Ces sources d’énergie soutiennent l’apparition de méthanogènes autotrophes, d’acétogènes et de bactéries ferrifères et sulfato-réductrices. Les bactéries sulfato-réductrices sont un constituant important du sous-sol terrestre et constituent souvent le groupe métabolique dominant où le sulfate est présent. Même lorsque le sulfate est limité, des bactéries sulfato-réductrices sont souvent détectées. Les accepteurs d’électrons ayant un potentiel de réduction plus élevé, tels que les nitrites, les nitrates et l’oxygène, sont généralement épuisés dans les profondeurs du sous-sol. Malgré une disponibilité limitée, le potentiel métabolique de réduction des nitrates a été identifié dans les eaux souterraines. La génération de sulfure, le produit métabolique de la réduction des sulfates, peut potentiellement compromettre la longévité des cartouches de cuivre utilisées pour stocker le combustible nucléaire usé. Il est donc crucial de comprendre les sources d’énergie qui alimentent la réduction des sulfates.
Des preuves d’un métabolisme dissimilatoire du soufre ont été recherchées en sondant le métagénome pour les gènes nécessaires à la réduction dissimilatoire du soufre; sulfate adénylyltransférase ( sat ), sous-unités alpha et bêta d’adénylylsulfate réductase ( aprAB ), sous-unités alpha et bêta dissimilatoires de sulfite réductase ( dsrAB ) et sulfite réductase dissimilatoire D ( dsrD ). La présence de dsrD a été considérée comme nécessaire pour qu’un MAG soit classé comme réducteur de sulfate, car ce gène est absent dans les micro-organismes qui utilisent la sulfite réductase à dissimilation inverse pour l’oxydation du soufre. Les gènes pour la dismutation du thiosulfate et du soufre élémentaire (polysulfures) sont annotés avec le même orthologue KEGG. Dans les génomes Desulfocapsa et Desulfurivibrio , la sous-unité catalytique de la thiosulfate réductase ( phsA )/polysulfure réductase ( psrA ) était adjacente à une ferrédoxine annotée comme sous-unité B de la tétrathionate réductase ( ttrB ), qui présente une grande similitude avec la même sous-unité. de polysulfure réductase ( psrB ). PsrAB nécessite également une sous-unité membranaire intégrale ( psrC ). Cette sous-unité appartient à la famille de type NrfD qui comprend les tétrathionate, DMSO, polysulfure et nitrite réductases. Les deux génomes codaient pour un gène annoté comme une sous-unité delta de la nitrate réductase adjacente au psrAB putatif , ainsi que pour une polysulfure réductase multiple ( nrfD ) ailleurs dans le génome. Ces gènes ( phsA/psrA, ttrB et nrfD ) peuvent être utilisés pour la réduction dissimilatoire du thiosulfate, du polysulfure ou du tétrathionate avec un donneur d’électrons tel que l’hydrogène. Dans des conditions standard, la dismutation du soufre est endergonique et nécessite que le sulfure soit réoxydé ou précipité avec des ions métalliques pour être thermodynamiquement favorable. Le fer (II) et le manganèse dissous sont tous deux détectés dans les eaux souterraines et pourraient constituer un puits pour les sulfures via les précipitations abiotiques. Nous proposons que le fer (II) dissous puisse être produit par l’activité métabolique du Rhodoferax . Ce MAG a codé la voie Mtr pour la réduction des métaux. Le cytochrome et la protéine de membrane externe associée au décaème (MtrB) ont été détectés dans le protéome, ce qui suggère que ce micro-organisme a la capacité de respirer des accepteurs d’électrons extracellulaires tels que le fer (III). En effet, dix MAG contenaient des gènes responsables du cycle oxydatif du soufre. L’activité métabolique des bactéries oxydant les sulfures a en outre été confirmée par la présence de peptides provenant de gènes oxydant les sulfures dans le protéome, indiquant que le sulfure est maintenu à de faibles concentrations dans ces eaux souterraines, au moins en partie par l’activité des sulfures. bactéries oxydantes. La première voie d’oxydation des sulfures considérée était la voie de réduction inverse des sulfites. De plus, le sulfite généré par la sulfite réductase à dissimilation inverse peut être oxydé en sulfate via l’adénylylsulfate réductase et la sulfate adénylyltransférase opérant dans le sens inverse ou par l’enzyme oxydant le sulfite cytoplasmique. Chez certaines espèces qui n’hébergent que du sqr , comme le Propionicimonas MAG, le sulfure peut être oxydé en soufre soit comme stratégie de détoxification du sulfure, soit comme stratégie d’assimilation du soufre. Desulfurivibrio alcaliphilus peut se développer en couplant l’oxydation des sulfures à une réduction dissimulatrice du nitrate en ammonium. Chez D. alcalinphilus , le seul gène connu associé à l’oxydation des sulfures détecté dans le génome était la sulfure:quinone oxydoréductase.
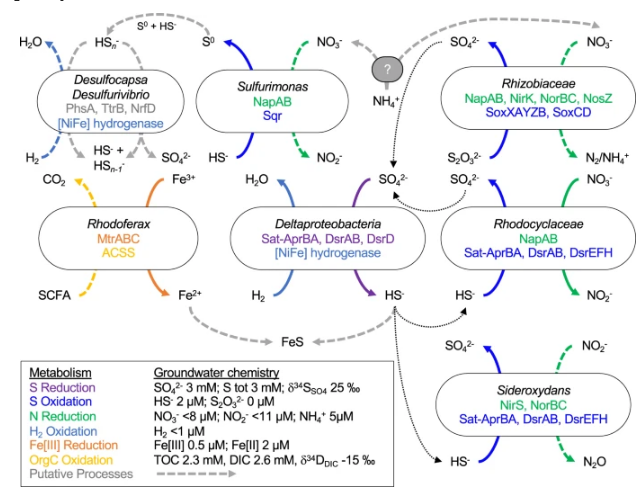
Les bactéries oxydantes des sulfures identifiées sont des anaérobies facultatifs. Leurs génomes contenaient des oxydases terminales pour la réduction de l’oxygène et des réductases pour la réduction des nitrates en N 2 , nitrite ou ammonium. La nitrate réductase ( napAB ) a été détectée dans le protéome des Rhodocyclaceae, ce qui suggère que les Rhodocyclaceae respirent activement du nitrate et de l’oxygène. Cette nitrate réductase périplasmique a une forte affinité pour le nitrate et peut être avantageuse dans un environnement limité en nitrate. La présence d’une oxydase de type cbb 3 de haute affinité suggère que les électrons provenant de l’oxydation de composés soufrés réduits pourraient être utilisés pour réduire l’oxygène dissous. Les Cbb 3 -oxydases peuvent également être exprimées lors de processus biologiques anaérobies tels que la fixation de l’azote et la dénitrification. Il a également été suggéré que l’oxydase de type cbb 3 élimine l’oxygène chez des espèces qui ne sont pas connues pour utiliser l’oxygène comme accepteur d’électrons, empêchant ainsi le stress oxydatif. La désulfocapsa exprime la nitrate réductase ( napA ) et la nitrite réductase ( nrfA ), qui catalysent ensemble la réduction du nitrate en ammoniac. De nombreuses bactéries sulfato-réductrices codent pour NapA et peuvent utiliser le nitrate comme accepteur d’électrons terminal alternatif. La nitrite réductase NrfAH se trouve couramment parmi les bactéries sulfato-réductrices et est proposée pour détoxifier les nitrites, supprimant l’inhibition de la réduction des sulfates par les nitrites. L’ammonium est détecté dans les eaux souterraines et pourrait éventuellement être oxydé par couplage à la réduction du sulfate, du fer ou du manganèse. Les composés azotés oxydés produits seraient alors rapidement utilisés et ne s’accumuleraient donc pas dans les eaux souterraines. Cependant, les enzymes nécessaires à cette oxydation anaérobie de l’ammonium ne sont pas bien comprises et nous n’avons pu déterminer ce métabolisme à partir des génomes récupérés. Il n’y avait également aucune preuve d’anammox.
Le carbone inorganique dissous est relativement élevé dans les eaux souterraines et peut constituer une source de carbone pour les micro-organismes autotrophes. Les enzymes clés de la voie réductrice de l’acétyl-CoA, la monoxyde de carbone déshydrogénase anaérobie ( cooS ) et l’acétyl-CoA synthase ( acsB ) ont été détectées dans toutes les MAG de Deltaproteobacteria. Deux voies de fixation du carbone ont été identifiées parmi les MAG oxydant les sulfures, la voie réductrice des pentoses phosphates et le cycle réducteur de l’acide tricarboxylique. Des peptides provenant d’enzymes clés de la voie réductrice des pentoses phosphates, la ribulose-1,5-biphosphate carboxylase (RubisCO) et la phosphoribulokinase, ont été détectés dans les protéomes des Rhodocyclaceae, Sideroxydans et Rhizobiaceae. Les sulfurimonas abritaient le cycle réducteur de l’acide tricarboxylique et l’enzyme clé ATP-citrate lyase ( aclAB ) a été détectée dans le protéome. On suppose donc que ces micro-organismes fixent le carbone inorganique. La métaprotéogénomique a révélé des oxydants de sulfures autotrophes actifs utilisant des espèces d’azote oxydées comme accepteurs d’électrons dans un environnement pauvre en sulfures. Le principal moteur du cycle du soufre est la réduction microbienne des sulfates, principalement par les Deltaprotéobactéries , qui produisent finalement du sulfure. es résultats ont des implications en termes de construction de stockage de déchets nucléaires sur ce site. La production de sulfure est préoccupante car elle peut corroder les cartouches de cuivre utilisées pour stocker le combustible nucléaire usé. L’activité des oxydants de sulfures pourrait donc constituer un puits bénéfique pour les sulfures dans les eaux souterraines d’Olkiluoto, ce qui pourrait limiter l’accumulation de sulfures après la fermeture du stockage. De même, l’oxydation des sulfures est vraisemblablement limitée par la disponibilité du nitrate mais reste néanmoins suffisante pour maintenir une faible concentration de sulfures dans les eaux souterraines.
Vie microbienne dans les roches granitiques profondes
https://academic.oup.com/femsre/article/20/3-4/399/516507?login=false
La roche granitique possède des aquifères qui traversent des failles et des systèmes de fractures simples ou multiples. Ils peuvent s’orienter dans n’importe quelle direction, verticalement ou horizontalement et, généralement, seules certaines parties des fractures des roches dures sont conductrices d’eau. Les parties restantes sont remplies de couches de minéraux précipités et d’argile et de gouge. L’échantillonnage des roches dures est difficile et le risque de contamination dû à l’intrusion de fluides de forage et de déblais dans les aquifères est évident. Une enquête récente sur le potentiel de contamination des forages dans le granit lors des opérations de forage, utilisant des méthodes moléculaires et de croissance, a montré que les micro-organismes prédominants dans l’équipement de forage étaient absents dans les eaux souterraines des forages forés. Le nombre total de bactéries trouvées dans les environnements granitiques souterrains varie de 10 3 à 10 7 cellules par ml d’eau souterraine, mais le nombre de micro-organismes cultivables est généralement beaucoup plus faible. Nous avons utilisé des techniques de culture avec taxonomie numérique pour l’identification de micro-organismes cultivables et la technique du gène ARNr 16S pour déterminer la diversité bactérienne dans les eaux souterraines granitiques. Des membres des genres Bacillus, Desulfovibrio, Desulfomicrobium, Eubacterium, Methanomicrobium, Pseudomonas, Serratia et Shewanella ont été trouvés. Plusieurs processus biogéochimiques dans les roches granitiques ont été mis en évidence dans lesquels les micro-organismes semblent jouer un rôle majeur. Un processus est la mobilisation des oxyhydroxydes de fer ferrique en phase solide en fer ferreux en phase liquide par des bactéries réductrices de fer avec du carbone organique comme donneur d’électrons. Un autre processus biogéochimique jugé important est la réduction du sulfate en sulfure par des bactéries réductrices de sulfate. Ils apparaissent fréquemment dans les aquifères granitiques en profondeur et semblent préférer une salinité modérée, environ 1 %. Lorsque les eaux souterraines riches en fer ferreux, en manganèse (II) et en composés soufrés réduits atteignent une atmosphère oxygénée telle qu’un tunnel ouvert, des gradients adaptés aux bactéries chimiolithotrophes se développent. Un troisième processus est la conversion du dioxyde de carbone en matière organique avec de l’hydrogène comme source d’énergie, éventuellement formé par radiolyse, réactions minérales ou par activité volcanique. Des résultats récents montrent que des méthanogènes autotrophes, des bactéries acétogènes et des méthanogènes acétoclasiques sont tous présents et actifs dans les roches granitiques profondes. Ces observations annoncent l’existence d’une biosphère profonde alimentée par l’hydrogène dans le substrat rocheux cristallin, indépendante de la photosynthèse. Si cette hypothèse est vraie, la vie pourrait avoir été présente et active dans les profondeurs de la terre depuis très longtemps, et on ne peut exclure que le lieu d’origine de la vie soit un environnement rocheux igné souterrain profond (probablement chaud avec une température élevée). pression) plutôt qu’un environnement de surface.
La faible teneur en Fe 2+ , par exemple , ne confère à l’eau qu’un faible pouvoir tampon rédox. Les bactéries réductrices de fer se sont révélées d’une importance biogéochimique majeure dans les roches granitiques. La stabilité redox inattendue du système étudié ne pouvait s’expliquer que par la mobilisation des oxy-hydroxydes de fer ferrique en phase solide en fer ferreux en phase liquide par des bactéries réductrices de fer avec du carbone organique comme donneur d’électrons.
Les bactéries acétogènes ont la capacité de faire réagir l’hydrogène avec le dioxyde de carbone pour former de l’acétate, produisant ainsi de l’ATP et réduisant l’énergie nécessaire au métabolisme. L’acétate produit peut être utilisé par des méthanogènes acétoclastiques, des bactéries réductrices de fer et de sulfate et d’autres micro-organismes hétérotrophes, constituant ainsi une voie de transformation du carbone inorganique en carbone organique avec de l’hydrogène comme réducteur.
https://www.nature.com/articles/s41467-019-10813-w?fromPaywallRec=true
Acidification
Processus contribuant à la diminution de la production de sulfure de diméthyle en réponse à l’acidification des océans dans les eaux subtropicales
L’influence de l’acidification des océans sur la production nette de diméthylsulfure (DMS), les concentrations de DMS intégrées dans le temps ont montré une relation inverse de −0,21 nmol DMS nmol −1 H + sur le gradient de concentration en H + de 8,8 à 23,3 nmol l −1 , ce qui équivaut à une plage de pCO 2 de 400 à 1 252 atm. Des diminutions proportionnellement similaires des concentrations de diméthylsulfoniopropionate (DMSP) dissous et particulaire ont été observées en relation avec l’augmentation de la concentration en H + entre les mésocosmes. La production nette réduite de DMSP avec une acidité accrue semble résulter d’une diminution de l’abondance d’une population de nanophytoplancton riche en DMSP. les taux spécifiques de production de DMS ont été significativement réduits à une concentration élevée en H + . Ces taux étaient étroitement corrélés aux taux de production nette de DMS, indiquant que la transformation du DMSP dissous en DMS par les bactéries était une composante majeure de la production de DMS. Il n’a pas été possible de déterminer si le catabolisme du DMSP était directement influencé par les concentrations en H + ou s’il s’agissait d’une réponse indirecte dans la composition de la communauté bactérienne associée à une disponibilité réduite de DMSP. Les altérations de l’émission de DMS par les vastes océans subtropicaux pourraient influencer la chimie atmosphérique et potentiellement le climat, sur une grande partie de la surface terrestre.
Les émissions de DMS contribuent à une réduction du forçage radiatif par un effet direct des aérosols et un effet indirect dû à la réflectivité des nuages. Le DMS est un produit principalement des communautés planctoniques microbiennes marines. Le précurseur du DMS, le diméthylsulfoniopropionate (DMSP), principalement synthétisé par les microalgues, est transformé en DMS ou DMSP dissous, par exsudation active, lyse cellulaire pendant la sénescence, lyse virale et broutage par le microzooplancton et le mésozooplancton. En raison de la consommation photochimique et biologique du DMS, seulement 2 à 10 % de la production de DMS est émise dans l’atmosphère. Le sort du DMS transformé biologiquement est mal connu, mais les produits les plus probables dans l’environnement pélagique sont le diméthylsulfoxyde (DMSO) et le sulfate. Une possibilité est que le métabolisme du DMS soit acheminé par le thiosulfate ou le tétrathionate, qui sont rapidement consommés pour produire du sulfate. Il existe de plus en plus de preuves expérimentales selon lesquelles les concentrations de DMS, et donc les émissions, sont susceptibles de changer en réponse à l’acidification des océans. Cela découle, en particulier, d’une série d’ expériences de perturbation de la pCO 2 dans des mésocosmes d’eau de mer naturels. Une tendance convaincante et cohérente de concentrations décroissantes de DMS avec une augmentation de la pCO 2 et des concentrations de protons [H + ] associées dans les eaux côtières.
Les relations entre [DMS] et le pH observées dans les expériences en mésocosme, en particulier l’expérience arctique, ont été utilisées comme base pour des prédictions modélisées de l’impact de l’acidification des océans sur les émissions mondiales de DMS et le climat. L’étude de Schwinger et al. (2017) a utilisé un modèle du système terrestre entièrement couplé pour démontrer une diminution potentielle de 31 % des émissions de DMS d’ici 2100, en supposant un niveau moyen, une réponse négative de la production de DMS au pH. Ce niveau de diminution des émissions de DMS pourrait contribuer à un réchauffement climatique supplémentaire de 0,3 K d’ici le tournant du siècle, en raison de la réduction de l’albédo des nuages et de l’effet direct des aérosols, car la charge en aérosols sulfatés est diminuée.
Il a été possible de distinguer plusieurs causes interdépendantes de la diminution de la production nette de DMS due à l’augmentation de l’acidité. Des niveaux d’acidité plus élevés ont diminué l’abondance des populations de nanophytoplancton qui dominaient la production de DMSPp. Cela a eu pour effet de diminuer la disponibilité de DMSPd. Les populations bactériennes ont pu répondre à cette diminution de la disponibilité du substrat en diminuant la vitesse spécifique à laquelle elles catabolisent le DMSPd, en particulier la vitesse à laquelle le DMSPd est clivé en DMS. Il reste à déterminer si ce changement dans le catabolisme du DMSP est le résultat d’une réponse physiologique des populations bactériennes existantes ou s’il reflète un changement dans la composition taxonomique.
https://www.frontiersin.org/journals/marine-science/articles/10.3389/fmars.2018.00245/full
https://www.sciencedirect.com/science/article/abs/pii/S0166445X20304380
https://www.sciencedirect.com/science/article/pii/S2095809921001442
https://www.sciencedirect.com/science/article/abs/pii/S0045653515005032
https://www.sciencedirect.com/science/article/abs/pii/S0045653504001596
https://www.sciencedirect.com/science/article/abs/pii/S0269749118316658
https://www.sciencedirect.com/science/article/abs/pii/S0048969717305971
https://www.sciencedirect.com/science/article/pii/S0043135423012976
https://www.sciencedirect.com/science/article/abs/pii/S0269749118316658
https://link.springer.com/article/10.1007/s11356-021-16483-w